Ассимиляция азота
Ассимиляция азота является одним из основных условий существования жизни на земле: организмы, не содержащие азота, неизвестны. В каждой живой клетке (см.) в составе протоплазмы, являющейся носительницей основных жизненных проявлений, важную роль играют белки (см.), содержащие 15-16% азота. Жизнь связана с их непрерывными глубокими превращениями. Обычно содержат азот и ферменты (см.), играющие первенствующую роль в физиологии обмена веществ. Наконец, в организмах встречается ряд других азотистых соединений. Все это достаточно характеризует физиологическую важность данного процесса и его большое практическое значение. Однако, вопрос этот представляет глубокий интерес и с более общей точки зрения. Азотистая пища и ее превращения у различных организмов очень разнообразны. На этой почве существует ряд поразительных соотношений, причем существование одних организмов оказывается тесно зависящим от обмена веществ остальных. Мало того, жизнь всех этих организмов близко связана с жизнью неорганической природы, ибо главная масса соединений азота, находящихся в земной коре, приходится на долю веществ органического происхождения. Все это дает один из наиболее ярких примеров того подвижного равновесия различных процессов, которое составляет основную черту столь поражающего нас всюду стройного порядка в природе.
Для питания человека и животных необходимы готовые белки. В составе обычной азотистой пищи на долю их приходится подавляющая по количеству и значению роль; содержание их в пище не может быть уменьшено ниже известного минимума. Свободный азот и его неорганические соединения не могут усвоиться животными. Усваивая белки, животный организм в то же время непрерывно разрушает их, превращая в более простые азотистые соединения; последние выделяются наружу в составе мочи и помета. Источником белковой пищи животных прямо (травоядные) или косвенно (плотоядные) являются растения. Таким образом, животное царство во всей его совокупности является непрерывным потребителем и разрушителем растительных белков. Что касается зеленых растений, то среди них есть немногочисленные представители, питающиеся подобно животным, готовыми азотистыми органическими соединениями: таковы насекомоядные, паразиты и сапрофиты (см.).
Но все это случаи уклоняющиеся, нетипичные. Подавляющая же масса зеленых растений в своем азотистом питании приурочена исключительно к солям азотной кислоты (нитратам), а также — аммиачным солям. Поглощая корнями эти неорганические соединения азота, растение перерабатывает их в азотистые органические вещества и созидает всю главную массу белков, встречаемых на земле. Всю жизнь зеленое растение, согласно общему характеру своего типа (см. растение), скопляет эти вещества в своем теле. Но жизненные процессы и здесь неразрывно связаны с обменом веществ, причем белки, как и в организме животных, подвергаются распаду. Однако судьба продуктов последнего в растениях совершенно иная. Здесь нет выделений, аналогичных обычным выделениям животных; азот, поглощенный растением, из организма последнего не выделяется, и продукты распада белков снова перерабатываются в белок. Растения, в общем, бедны азотом, содержа его в среднем не более 11/2% своего сухого вещества, то есть лишенного воды (см. агрономический анализ). Однако в сумме запросы на нитраты, предъявляемые растительностью к почве, велики. Каждая жатва уносит в среднем до 50 килограммов азота нитратов на гектар. В почве же последних обычно очень мало, аммиака — еще меньше. Поэтому голодание на азот является самой обычной причиной неурожаев, и вопрос об азотистых удобрениях исстари стал одним из основных вопросов техники и экономики земледелия. Источником азота для зеленого растения является отчасти еще и аммиак атмосферы, который может усвояться листьями. Однако, его мало, и прибыль азота в растениях при усвоении его из этого источника практически ничтожна. Наконец, неисчерпаемый источник азота зеленые растения могли бы иметь в свободном азоте воздуха. Однако, сами по себе усвоить его они не могут (Буссенго, 1854). В гораздо больших количествах, чем нитраты и аммиак, корни растений встречают в почве азотистые органические вещества отбросов и трупов (будут ли они естественно накоплены в ней, или внесены с навозным удобрением). Некоторые из этих веществ в искусственных условиях могут служить азотистой пищей для зеленого растения. Однако, в естественной обстановке эти вещества быстро подвергаются гниению (см.), делаясь добычей целого ряда грибов и, главным образом, бактерий, массами заселяющих почву. Эти микробы поглощают азотистые органические вещества, перерабатывают их в более простые соединения и, подобно животным, обратно выделяют эти продукты в окружающую среду. Тип обмена веществ разных бактерий обнаруживает глубокие различия; так что вещества, являющиеся отбросами деятельности одних, служат пищей для других. Благодаря этому, микробы сменяют друг друга, обусловливая ряд как бы налагающихся один на другой процессов. Сначала одни из бактерий нападают на белки и отщепляют часть их азота в виде аммиака, другую же, большую часть оставляют в виде азотистых органических соединений, более простых, чем белок. Эти последние делаются добычей новых бактерий, вырабатывающих еще более простые продукты и аммиак. Той же судьбе подвергаются и азотистые небелковые отбросы животного организма; среди них выделяемая в больших количествах мочевина особыми микробами (уробактерии) перерабатывается в углеаммонийную соль. Все эти процессы, объединяемые общим названием аммонизации, переводят, в конце концов, весь органический азот в аммонийные соли. Эти последние, однако, быстро подвергаются так называемой нитрификации, то есть перерабатываются специфическими бактериями сначала в соли азотистой кислоты, а затем новыми бактериями в соли азотной кислоты. Таким образом, органический азот оказывается нацело минерализованным, то есть переведенным в неорганические соединения, и лишь после того потребляется зеленым растением. Нитрифицируется обычно и вносимая в виде удобрения серно-аммиачная соль. Нитрификация идет и в почвах, и в водах повсеместно и непрерывно, достигая в жаркое время года и в теплом климате своего наибольшего напряжения. Этим биологическим процессам обязана своим происхождением почти вся масса нитратов на земле; лишь ничтожное количество их, как увидим, образуется чисто химическим путем. Тех же бактерий должны мы признать крупным фактором и в геологическом прошлом земли: им, несомненно, обязаны своим происхождением колоссальные залежи селитры в Чили. Те же бактерии еще недавно играли крупную роль и в техническом получении нитратов. Искусственно создаваемые из земли, золы, хвороста и всевозможных органических отбросов (навоза и проч.), очаги работы микробов-аммонизаторов и нитрификаторов, носившие название селитряниц, являлись важным фактором в области военного дела (в целях получения пороха); они долгое время составляли предмет правительственных забот, пока не найдены были другие источники нитратов и другие взрывчатые смеси. Образовать белки, пользуясь неорганическими соединениями азота (главным образом аммиаком), могут и некоторые грибы и бактерии; но в целом, сравнительно с синтезом белков зеленым растением, процесс этот доставляет лишь ничтожно малые количества этих веществ.
Если подвести итоги сказанному, то мы получим картину без остановочного перехода азота из одного состояния в другое; и весь этот круговорот поддерживается обменом веществ различных организмов. Не трудно заключить, что для поддержания существующего порядка вещей на земле, не только не может быть устранено ни одно из звеньев этого круговорота, но и с количественной стороны все они должны быть взаимно уравновешены. Число людей и животных тесно зависит от общего количества зеленых растений. О той же количественной зависимости благосостояния животных и зеленых растений от микробов почвы должен говорить нам любой случай, когда голодание посева на азот грозит неурожаем, предвещающим голод и тяжелые экономические последствия для населения. Наконец, в отношении азотистого питания растения — ряд таких культурных приемов, как удобрение и обработка почв является, в сущности, вмешательством в круговорот азота, с целью повысить интенсивность работы отдельных его звеньев (путем ли внесения веществ, потребляемых почвенными микробами — навоз — или вырабатываемых последними — нитраты, или путем создания условий, благоприятных для их деятельности — разрыхление и т.п.).
При ближайшем исследовании, однако, обнаружилось, что круговорот азота не так прост: в нем нашлись крупные осложнения. Первое из них обусловливается чрезвычайной подвижностью нитратов, столь ценных для зеленого растения. Когда количество почвенной влаги от дождей или снега повышается, она начинает просачиваться и стекать в ручьи и реки, последними же уносится в моря. Другие вещества при этом задерживаются из раствора, благодаря так называемой поглотительной способности почв; нитраты же лишены этого свойства и беспрепятственно вымываются. Подсчет этого процесса для больших рек дает такие высокие цифры суточного выноса селитры в море, как 238 000 кг (Сена в среднюю воду), или 1 миллион кг (Нил). Потери эти являются, однако, лишь местными: селитра только перемещается в море. Здесь ее азот повторяет только что рассмотренный цикл, переходя от накопляющих белки водорослей к пожирающим их морским животным, а с отбросами последних и трупами — к микробам-минерализаторам. Однако, вследствие затрудненного доступа кислорода в водные толщи, нитрификация идет здесь слабо, и аммиак скопляется в морской воде. Отсюда он постоянно выделяется, и ветры несут его на материки. Здесь он падает на землю с осадками и отчасти усвояется листвой растений. Подсчеты количеств аммиака, приносимых таким путем на землю, для южных стран дали цифры высокие (до 50 килограммов аммиака в год на гектар — Франция), для других — значительно более низкие. Эта так называемая циркуляция аммиака (Шлёзинг, 1875) является противовесом вымыванию селитры.
Гораздо более грозным представляется другой факт: в круговороте азота часть его постоянно выделяется в свободном виде. Таким образом, общий запас азотистых соединений постоянно убывает, а это, очевидно, угрожает, в конце концов, самому существованию жизни на земле. Известны бактерии, которые при затрудненном доступе кислорода воздуха и обильном питании органическими веществами, способны отнимать кислород от нитратов, разрушая их с образованием свободного азота. Возбудители этой так называемой денитрификации крайне распространены в навозе, почвах, грязных водах, морях и проч. Процессы эти могут выделить до 90% азота из нитратов и идут очень энергично. Наконец, свободный азот выделяется при горении азотистых органических соединений. Каковы процессы, компенсирующие эти потери, — долго оставалось загадочным. Давно было известно, что при действии ряда электрических искр происходит соединение азота с кислородом; благодаря этому, гроза доставляет почве с дождевой водой небольшие количества азотной кислоты. Далее, под влиянием тихого электрического разряда некоторые органические вещества фиксируют азот. Однако, все эти процессы количественно чрезвычайно слабы и неспособны компенсировать вышеуказанных потерь связанного азота. Между тем, еще прежде, чем во всей полноте выяснились все эти обстоятельства, учет массы азота, циркулирующей в хозяйстве, показал, что количество связанного азота, бывшего в почве и внесенного удобрением, дождями и проч., в сумме значительно ниже количества его, взятого с этой площади за период полного севооборота урожаями. Очевидно, какой-то процесс обогащал почву на счет не входившего в учет свободного азота.
Далее, среди культурных растений еще римляне отметили бобовые — горох, лупин, вика и проч., — как ведущие себя совершенно своеобразно. Другие растения истощали почву; между тем, культура бобовых не только не вызывала этого, но действовала даже благоприятно на последующие посевы, как бы удобряла почву. Это повело к выработке так называемого зеленого удобрения, запахиванию посева бобовых в период цветения с целью повысить плодородие почвы. Далее, бобовые, несмотря на то, что они содержат гораздо больше азота, чем другие растения, роскошно развиваются на почвах, совершенно им бедных. Мало того, после культуры злаков содержание азота в почве падает, после бобовых — возрастает. В виду этого возникло деление растений на азотособирателей (см.) — бобовые — и азоторасточителей — злаки и проч. В среде, лишенной азота, при постепенном обогащении ее (до известного предела) нитратами, на 1 весовую единицу последних злаки отвечали повышением урожая (см. рисунок 1 таблицы) приблизительно на 100 весовых единиц сухого вещества. Бобовые давали прирост последнего без всякой зависимости от нитратов (см. рисунок 2). Вся капризность этого явления, однако, исчезла, как только было обращено внимание на одно обстоятельство. На корнях бобовых давно было известно присутствие многочисленных вздутий, — так называемых клубешков (см. рисунок 6). В клетках последних известно было присутствие бактерий. Раньше это явление считалось болезненным. Теперь же найден был замечательный параллелизм его с особенностями азотистого питания. Стоило стерилизацией среды, посуды и семян устранить бактерий, а следовательно и помешать образованию клубешков, — и бобовые начинали вести себя в отношении к нитратам подобно злакам и оказывались неспособными усваивать свободный азот (см. рисунок 3). Стоило заразить среду клубешковыми (но не какими-нибудь вообще почвенными) бактериями, — появлялись клубешки, а с ними и способность усваивать свободный азот (Гелльригель и Вильфарт, 1887). Бактерии эти, получившие название Bacillus radicicola, в искусственных средах, богатых углеводами, оказались и вне растения способными усваивать азот. Здесь мы имеем интереснейший пример сожития на началах обоюдной пользы, так называемого симбиоза (см.). Весной бактерии из почвы проникают в разных местах в поверхностные клетки молодых корешков, где и размножаются, образуя слизистые скопления (см. рисунок 9). Отсюда они постепенно внедряются в окружающие клетки, давая чисто местные очаги заражения. Пораженные клетки не гибнут, а, наоборот, разрастаются и размножаются. Это вызывает припухлость, превращающуюся затем в клубешок. В нем бактерии заселяют крупные клетки его внутренней массы (см. рисунки 7 и 8). Последняя окружена тканью, содержащей большие количества крахмала. Клубешки развиваются быстро и первое время угнетают растение-хозяина. Бактерии явно живут на счет бобового, пользуясь от него необходимыми для них безазотистыми органическими веществами (крахмал). Последние зеленое растение синтезирует в изобилии из воды и углекислоты, недостатка в которых обычно нет. Вскоре картина отчасти меняется. Защитные слизистые массы начинают исчезать, и бактерии обнаруживают тогда признаки угнетения (см. рисунок 10 b); вместе е тем, растение-хозяин оправляется и начинает быстро развиваться. Клубешок, бывший до тех пор очень богатым сухим веществом и азотом, начинает терять и то, и другое. Очевидно, растение-хозяин губит бактерий и всасывает недостающие ему синтезированные бактериями азотистые вещества. Осенью, при сгнивании корней, уцелевшие бактерии выходят в почву. Органический же азот, скопленный в остатках жатвы или в зеленом удобрении, минерализуется и, в конце концов, (косвенно) обогащает почву нитратами. Так разъяснилась историческая загадка бобовых и их роль в севообороте. Это важное открытие в области теории сейчас же привело и к совершенно новому принципу в области практики земледелия, — к так называемому бактериальному удобрению: внесению вместе с семенами бобового культуры его клубешковых бактерий (так называемый нитрагин). Присутствие клубешков и способность усвоять свободный азот свойственны не только всем бобовым, но и отдельным представителям немногих других семейств, каковы, например, ольха (см. рисунки 4 и 5), лох и облепиха. Аналогичное явление наблюдается и среди низших водорослей, покрывающих иногда густым зеленым налетом поверхность земли и в чаще посева, и на паровом поле, и в естественной дерновине. В этих условиях почвы связывают свободный азот; мало того, будучи первоначально совершенно бедны им, они оказываются при этом способными доставлять хорошее азотистое питание не имеющим клубешков растениям. Деятелями и здесь оказались какие-то сожительствующие с водорослями бактерии. Здесь опять мы имеем случай симбиоза: отдавая водорослям связанный азот, полученный из недоступного для них источника, бактерии получают безазотистые органические вещества, которые сами синтезировать не могут и которые, наоборот, легко накопляются водорослями.
Еще более крупную роль в процессе ассимиляции элементарного азота играют свободно живущие в почве и водах бактерии. Первым был найден крупный бацилл — Clostridium Pastorianum (Виноградский, 1893). Ведя масляно-кислое брожение сахаристых веществ в условиях отсутствия кислорода и каких-либо соединений азота, микроб этот энергично ассимилирует свободный азот, переводя его в азотистые органические вещества. Микроб этот, будучи анаэробом, в то же время может развиваться лишь при условии легкого доступа свободного азота. Эти, казалось бы, трудно реализуемые в природе условия создаются опять-таки симбиозом: Clostridium Pastorianum живет с двумя бактериями - спутниками, аэробами, свободного азота не усваивающими.
Плотно окружая своих сожителей, они не мешают его доступу к последним; поглощая же атмосферный кислород, создают необходимые для них условия анаэробиоза. В свою очередь спутники, очевидно, получают связанный азот (существуют и другие выгоды этого сожития). Микроб этот распространен в самых разнообразных почвах и в морской воде. Существуют и аэробные виды, усваивающие свободный азот. Один из них, так называемый Azotobacter, широко распространен в почвах, в плавучей флоре поверхности морей, на водорослях, в сточных водах и проч. Сбраживая различные безазотистые органические вещества, он образует массу буреющей слизи и усваивает свободный азот гораздо интенсивнее, чем Clostridium Pastorianum. Наконец, усваивают азот (хотя и слабее предыдущих) некоторые другие, менее изученные микробы.
В деятельности всех этих организмов процессы, освобождающие азот из его соединений, находят достаточно мощный противовес.
Е. Вотчал.
Ассимиляция азота.
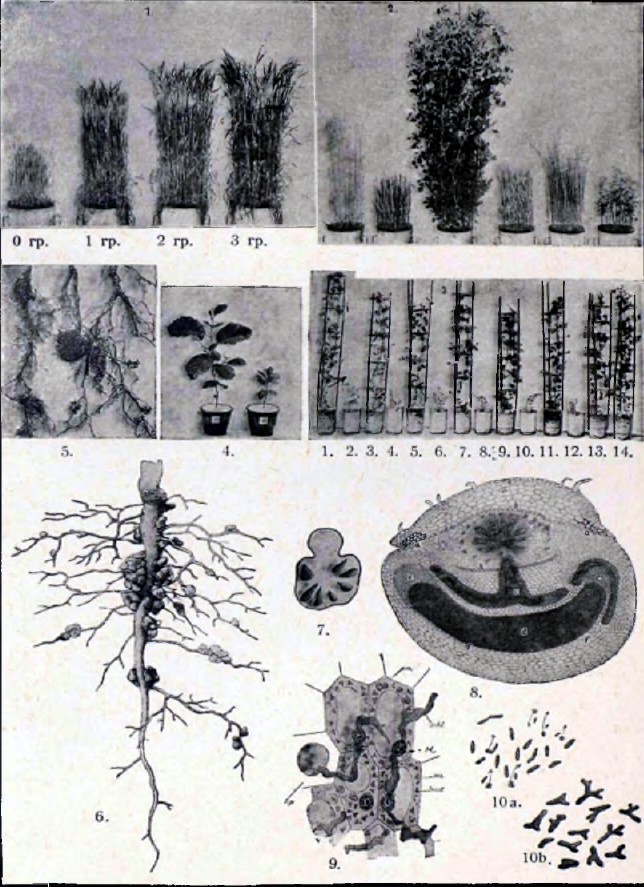
ОБЪЯСНЕНИЕ ТАБЛИЦЫ К СТАТЬЕ «Ассимиляция азота».
Рисунок 1. Развитие ячменя в зависимости (при прочих равных условиях) от различных количеств (0 грамм, 1 г, 2 г, 3 г) азота селитры в почве.
Рисунок 2. Развитие ржи, овса, гороха, пшеницы, льна и гречихи на почве, почти совершенно лишенной нитратов, но содержащей в достаточном количестве остальные необходимые питательные вещества.
Рисунок 3. Параллельные культуры гороха в нестерилизованной (№№1, 3, 5, 7, 9, 11, 13 и 14) и стерилизованной (№№2, 4, 6, 8, 10, 12), лишенной связанного азота почве, иллюстрирующие потерю способности усваивать свободный азот и развиваться независимо от нитратов после стерилизации почвы (из работы Гелльригеля 1887 года).
Рисунок 4. 11-месячные ольхи, развившиеся в почве, лишенной связанного азота — не стерилизованной (сосуд №1) и стерилизованной (сосуд №2).
Рисунок 5. Клубешки на корнях ольхи.
Рисунок 6. Корень желтого лупина с клубешками.
Рисунок 7. Поперечный разрез корня лупина в том месте, где образовался крупный клубешок. Часть, обращенная к верхней стороне таблицы, — разрез самого корня. Более широкая часть, обращенная вниз, — разрез клубешка, в котором находятся (ясно различимые простым глазом) расположенные гнездами участки нежной, обычно розовеющей при разрезе ткани, занятой бактериями.
Рисунок 8. Микроскопический вид разреза через тонкий корень лупина, несущий молоденький клубешок. Верхняя половина рисунка — разрез корня; нижняя (за aha) — разрез клубешка; zb — ткань коры корня, со стороны клубешка разорванная выступившим из нее клубешком; g — проводящий вещества по растению сосудистый пучок, посылающий ветки (с) в клубешок; d — одно из гнезд внутренней нежной ткани клубешка (отмеченной на рисунке темной штриховкой), заселенной Bacillus radicicola. На концах (ее) ткань эта еще продолжает разрастаться; f — более плотная ткань коры клубешка, окружающая ткань, занятую бактериями, служащая обычно местом отложения крахмала.
Рисунок 9. Микроскопический вид части разреза через внутреннюю, заселенную бактериями часть клубешка Lathyrus silvestris. В протоплазме (hp) клеток видны переходящие из одной клетки в другую слизистые нитевидные скопления Bacillus radicicola (schl), направляющиеся обычно к клеточным ядрам (k); в той же протоплазме видны ее мелкие зернышки, так называемые микрозомы (mi), зерна крахмала (am) и отдельные освободившиеся из слизи бактерии (bact), обнаруживающие ветвления, вздутия и прочие признаки угнетения.
Рисунок 10. Bacillus radicicola: а) нормальные бактерии, выделенные в искусственной культуре из клубешков гороха. Некоторые — движутся в направлении, указанном стрелками; b) бактерии из клеток клубешков того же гороха, обнаруживающие признаки угнетения (так называемые «бактероиды»).
| Номер тома | 4 |
| Номер (-а) страницы | 90 |