Электрофизиология
Электрофизиология, отдел физиологии, посвященный изучению электрических явлений, развивающихся и протекающих в животном организме; электрофизиологию часто называют, поэтому, учением о животном электричестве.
История развития учения о животном электричестве. Вопрос о том, является ли тело животного источником электрических сил, возник очень рано, еще в ту пору, когда сама физика электричества находилась в младенческом состоянии, когда все знания об электричестве ограничивались лишь первоначальными понятиями о заряде и разряде тел. Мысль о животном электричестве родилась раньше, чем возникло учение об электрическом токе, — даже больше: именно на почве неустанных попыток обнаружить животное электричество и создана была впервые случайно та обстановка, при которой осуществились условия для получения электрического тока, и первым его показателем был живой объект — лягушечья лапка. История животного электричества и история открытия электрического тока сплетены в первых их стадиях теснейшим образом. Здесь было много случайного, много счастливых моментов и наряду с этим много недоразумений и взаимного непонимания физиков и физиологов, пока не наметились, наконец, разные их пути. Мы укажем лишь главные эпизоды в этой интересной главе истории естествознания, которая начинается наблюдениями Луиджи Гальвани (см.), подробно изложенными в ст. гальванизм (см. XII, 419 сл.).
Из многочисленных опытов Гальвани остановимся здесь на одном, который имел громадный успех и всюду с большим увлечением повторялся всеми, кого интересовало животное электричество. На стеклянную пластинку кладут нервно-мышечный препарат, берут в руки две металлические палочки и прикладывают их концами одну к мышце, другую к нерву; если теперь соединить другие, свободные концы палочки друг с другом, то получается сокращение мышц. Опыт идет особенно хорошо, если палочки приготовлены из разных металлов, например, из меди и цинка. Видеть в указанном опыте проявление электрических сил можно было, по толкованию Гальвани, потому, что только соединение проводниками электричества мышцы и нерва давало сокращения; если вместо металлов взять для соединения непроводники — сургуч, стекло и др., то сокращений нет. Так как Гальвани считал, что в указанной цепи металлы были лишь проводниками, то для него источником электричества были животные части. Описанный опыт считался поэтому на первых порах, в самом деле, строгим доказательством существования животного электричества. Но вскоре обстоятельства изменились. На сцену выступает знаменитый современник, впоследствии противник Гальвани, профессор физики Александр Вольта (см.). Повторяя опыты Гальвани, он скоро увидел некоторые его промахи. Между прочим, он сделал один простой и на первый взгляд наивный опыт, имевший громадные научные последствия. Вольта повторил опыт Гальвани не на лягушечьем препарате, а на мышце человека, мышце, легко доступной и покрытой лишь тонким слоем эпителия, — на языке. Вольта приложил два кусочка разных металлов к двум точкам языка: при замыкании этой цепи он не мог отметить движений языка. Он был, однако, чрезвычайно поражен тем, что при замыкании описанной цепи он почувствовал особый вкус во рту: кислый и щелочной. Существенно здесь было то, что вкус не был приурочен к определенному месту языка, а зависел от металла, приложенного к языку.
Вольта заключил отсюда, что металлы в этом опыте являются не только проводниками электричества, а скорее — наоборот: из металлической цепи устремляется в живую часть цепи нечто, что дает вкусовое ощущение. С другой стороны, Вольта оценил тот знаменательный факт, что ощущение на языке длится в течение всего времени, пока цепь замкнута. Отсюда он пришел к заключению, что здесь мы имеем дело не с разрядом электричества, а с непрерывным течением его. Таким образом, Вольта впервые высказал мысль об электрическом токе. Животные части цепи оказались в понимании Вольты совершенно пассивными звеньями. Вольта заменил их проводящими жидкостями, построил дальше свой, теперь так называемый вольтов столб (см. XII, 424/26) и отрицал всякое значение опытов Гальвани как опоры для доказательства животного электричества. Гальвани испытал все превратности судьбы, но он не сдавался. Чтобы восстановить опороченную роль лягушечьих лапок как источника животного электричества, он стал теперь делать свои опыты так, чтобы не могло быть подозрения об участии в них металлов, и дошел, наконец, до такой формы опыта, которую мы и теперь с полным убеждением считаем доказательством биоэлектричества. Гальвани отпрепарировал мышцу с ее нервом; он набрасывал стеклянной или костяной палочкой нерв на мышцу и замечал, что в момент прикосновения нерва с мышцей последняя сокращалась. Это знаменитые опыты Гальвани — «опыты сокращения без металлов», из-за которых Гальвани по праву считается отцом электрофизиологии. Дальше этого Гальвани, однако, не мог пойти. Нетрудно усмотреть, что в этих последних опытах мышца играла двойственную роль: она была и источником электричества, и в то же время его показателем, и это обстоятельство не давало возможности в точности проанализировать установленное им явление. Опыты сокращения без металлов были, поэтому, на долгое время забыты, и только когда Нобили построил мультипликатор с астатическими магнитными стрелками (см. XII, 452), отличающийся большой чувствительностью, явилась мысль ввести в одну и ту же цепь и гальванометр, и лягушечью мышцу, чтобы решить, какой показатель чувствительнее к электрическим токам, живой или мертвый. Включение в одну и ту же цепь гальванометра и мышцы и было той постановкой опыта, которая должна была заменить «опыты сокращения без металлов». Мышца освобождалась теперь от роли показателя электрического тока и исследовалась исключительно как источник электричества. После целого ряда исследований самого Нобили, Гумбольдта, Маттеуччи весь вопрос о животном электричестве переходит на много лет в руки берлинского физиолога и прекрасного физика Э. Дюбуа-Реймона (см.). Этому ученому мы, прежде всего, обязаны выработкой весьма совершенной электрофизиологической методики, которою мы в значительной мере пользуемся еще и в настоящее время: его ключ, его санный аппарат, особенно его неполяризующиеся электроды и др. являются обыкновенным инвентарем современной физиологической лаборатории. Дюбуа-Реймон установил, прежде всего, так называемый «ток покоя». По его терминологии всякая вырезанная из тела мышца, например, лягушечья портняжная мышца, построенная из параллельных мышечных волокон, имеет так называемую внешнюю продольную поверхность, два поперечных разреза на концах и экваториальную линию посередине. Если различные точки такой мышцы соединить с гальванометром, то можно убедиться, что мышца служит источником электрических токов. Направление и величина токов различны в зависимости от выбранных для отведения в гальванометр точек мышцы. Дюбуа-Реймон отметил в этом отношении такое правило: от симметричных точек мышцы, например, от двух точек обоих поперечных разрезов или от двух точек поверхности, равно отстоящих от обоих поперечных разрезов, нельзя отвести тока; стрелка гальванометра остается на нуле. Не симметричные же точки дают ток, и чем менее они симметричны, тем большая между ними разность потенциалов. Поэтому точка экватора и центральная точка поперечного разреза дают максимальную разность потенциалов. В этих случаях мышца является как бы гальваническим элементом, причем продольная поверхность мышцы служит положительным полюсом этого элемента, а поперечный разрез — отрицательным. Дюбуа-Реймон применил способ Погендорфа для измерения электровозбудительной силы и нашел, что для лягушечьей мышцы величина ее колеблется от 30 до 80 милливольтов. Вырезанный из тела нерв, например, седалищный нерв лягушки, тоже подчиняющийся указанным законностям, обнаруживает электровозбудительную силу от 10 до 30 милливольтов. Таким образом, было раз и навсегда установлено, что мышца и нерв в описанных условиях служат источниками токов, и именно так называемых «токов покоя», ибо они дают токи, сами находясь при этом в покойном состоянии. Дюбуа-Реймон установил дальше явление исключительной важности: он показал, как изменяются токи покоя, когда мышца или нерв, от которых отводят ток, приходят в возбужденное состояние под влиянием быстро следующих друг за другом одиночных раздражений. Если мышцу, дающую ток, привести таким образом в состояние тетануса (см. II, 616/17), то ее ток покоя становится меньше, стрелка гальванометра устремляется в направлении к нулю, то есть идет в сторону, обратную первоначальному отклонению, — это так называемое «отрицательное колебание тока покоя». О том, что мышца возбуждена, мы можем, конечно, судить и, помимо отрицательного колебания, по ее сокращению. Судить же при помощи глаза, возбужден ли нерв, мы не в состоянии, ибо никаких видимых для глаза изменений нерв при своем возбуждении не обнаруживает. В гальванометре мы приобретаем, таким образом, инструмент, который открывает нам возможность судить о возбуждении нерва даже в том случае, если последний не соединен с мышцей: отрицательное колебание тока покоя нерва есть свидетельство его возбужденного состояния. Конечно, естественно возникает вопрос, как смотреть на отрицательное колебание: есть ли это акт прерывистый, колебательный, или постоянно текущий. Дюбуа-Реймон решил этот вопрос, использовавши так называемое явление вторичного сокращения. Этот опыт, описанный впервые Карло Маттеуччи (итальянский физик, 1811-1868, профессор в Болонье, Равенне, Пизе, написавший «Lezioni di fisica», 1841, и «Lezioni sui fenomeni fisico-chimici dei corpi viventi», 1844), состоит в следующем: из двух нервно-мышечных препаратов икроножной мышцы с седалищным нервом (см. рис. 1) накладывают нерв первого препарата на мышцу второго, раздражают тетанически нерв второго препарата в месте а и вызывают, таким образом, тетаническое сокращение мышцы этого второго препарата.
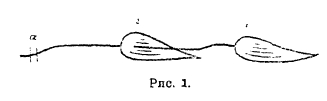
Рис. 1.
Но поразительно при этом то, что в тетаническое сокращение впадает и мышца первого препарата. Объяснение, данное этому замечательному явлению Дюбуа-Реймоном, сводится к тому, что отрицательное колебание токов в тетанизированной второй мышце есть явление прерывистое, причем каждому отдельному, одиночному раздражающему стимулу соответствует и определенное одиночное уменьшение тока покоя. Гальванометр, однако, вследствие своей инертности, не в состоянии обнаружить все отдельные моменты, из которых слагается отрицательное колебание, и поэтому дает лишь суммарное отклонение в виде отрицательного колебания. Если же на мышцу мы кладем нерв, то он, как образование, быстрее реагирующее на проходящие через него прерывистые мышечные токи тетанически сокращающейся мышцы, вызывает тетанус в той мышце, которую он сам иннервирует. Понятно поэтому, что в течение всего времени, пока длится раздражение второй мышцы и, следовательно, в течение всего времени, пока длится ее отрицательное колебание, первая мышца будет в состоянии тетануса.
Гипотеза Дюбуа-Реймона. Представленные данные о токах покоя и отрицательного колебания укладывались хорошо в предложенную Дюбуа-Реймоном гипотезу о биполярных и периполярных электрических молекулах, из которых, согласно этой гипотезе, построены возбудимые ткани. Каждая молекула имеет в своей средней части положительную поверхность, а в обеих полярных частях — отрицательную поверхность. Возбудимая ткань, сложенная из таких биполярных молекул, должна, как правильно доказывал Дюбуа-Реймон, давать такие результаты распределения потенциалов и направления отводимых токов, какие, в самом деле, наблюдаются на мышце. Модель объясняла без всякого труда и тот факт, что повторные поперечные разрезы давали в результате опять образование с отрицательным поперечным разрезом и положительною продольною поверхностью. Для объяснения отрицательного колебания оказалось необходимым приписать молекулам новое свойство — вращение: при раздражении мышцы ее ток покоя вследствие этого вращения должен уменьшиться. Нетрудно усмотреть, что по теории Дюбуа-Реймона токи покоя должны были иметь место и в неповрежденной мышце, ибо и в ней принцип расположения молекул оставался таким же, как и в поврежденной. Это и признавал Дюбуа-Реймон. Он отличал даже поперечный искусственный разрез от естественного разреза, причем под последним понималась сумма концевых поверхностей мышечных волокон, упиравшихся в сухожилие. «Предсуществующие» в мышце молекулы должны были при отведении от сухожилия (как бы приложенного к естественному поперечному разрезу) и от поверхности мышцы давать токи покоя неповрежденной мышцы. Хотя токи покоя неповрежденной мышцы и входили как принципиально необходимое звено в теоретическое построение Дюбуа-Реймона, тем не менее, он сам неоднократно испытывал затруднения при демонстрации этих токов. Токи покоя неповрежденной мышцы большей частью были слабы, скоропреходящи или, наоборот, лишь постепенно развивались, иногда же совсем отсутствовали. Дюбуа-Реймону пришлось по этому поводу предположить существование особенного «противозаконного» слоя («парэлектрономия») молекул на границе сухожилий с мышцей, мешающего току покоя обнаружить себя. Однако, все эти поправки свидетельствовали лишь о насилии теории над фактами, и, как всегда, факт оказался победителем.
Альтерационная теория Германа. Людимар Герман (1838-1914, с 1884 года профессор в Кенигсберге, написал: «Lehrbuch der Physiologie des Menschen», 14-oe изд. 1900. «Untersuchungen zur Physiologie der Muskeln und Nerven», 1867-68, и др.), знаменитый современник Дюбуа-Реймона, отчасти его ученик, а затем сильнейший противник, тщательно обставленными опытами доказал, что токи покоя в естественных условиях, то есть на неповрежденных тканях, не могут быть обнаружены; малейшее же повреждение их, ничтожная царапина на мышце, воздействие какого-либо химического вещества сейчас же дают начало токам покоя. Герман показал, что знаменитый опыт Гальвани сокращения без металлов, то есть сокращения мышцы при набрасывании на нее ее же нерва, удается лишь тогда, если набрасываемый нерв попадает на поврежденное и неповрежденное место мышцы; только в этом случае, вследствие образования потенциала у места надреза, могут возникнуть токи покоя, выравнивающиеся через нерв и раздражающие его. По Герману, нет токов покоя без повреждения. Если же это так, то вряд ли можно принимать существование биполярных электрических молекул в неповрежденной мышце, как этого требовала теория Дюбуа-Реймона. Если же, дальше, нет молекул в цельной мышце, то нельзя допустить, чтобы они могли образоваться в момент повреждения мышцы. Теория Дюбуа-Реймона должна быть, поэтому, оставлена. По представлению Германа, в месте повреждения возникает какое-то, ближе невыясненное, изменение в живой мышечной ткани, связанное с образованием кислоты. Это изменение, «альтерация», ведет к появлению на границе между живой, совершенно неизмененной тканью, и измененной, отмирающей тканью — потенциала, причем роль отрицательного полюса играет измененная вследствие повреждения часть, а роль положительного полюса — неизмененная часть мышцы. Все правила Дюбуа-Реймона о распределении сильных и слабых потенциалов на мышце с поперечными разрезами можно без труда вывести и из модели Германа. Целый ряд фактов подтверждает точку зрения Германа. Так, например, скелетная мышца с поперечным разрезом через все ее волокна дает ток покоя до тех пор, пока все волокна не погибнут; если же нанести повреждение на поверхности желудочка лягушечьего сердца с его короткими, друг с другом связанными мышечными волокнами, то от пораненного места и от нетронутой поверхности можно отвести ток покоя, который, однако, через короткое время исчезнет, как только поврежденные волокна погибнут, и вместо прежней границы между живым и отмирающим появится граница между живым и мертвым. Ток покоя вследствие поперечного разреза на нерве держится тоже недолго: как только отмирание дойдет до ближайших узлов Ранвье (XX, 247/48), то прекращается и ток покоя. Большое преимущество теории Германа в том, что в ее рамки совершенно естественно укладывается и объяснение явлений отрицательного колебания. Герман принимает, что существенным для процесса возбуждения в какой бы то ни было возбудимой ткани является то обстоятельство, что на границе между возбужденным и покойным участком появляется разность потенциалов; при этом возбужденное место играет роль отрицательного полюса, а невозбужденное место - положительного. Если поэтому у нас мышца с поперечным разрезом и электроды приложены так, что ток проходит через гальванометр от продольной поверхности к поперечному разрезу, то при раздражении такой мышцы возбуждение охватит и ту точку на продольной поверхности, на которой находится электрод. Вследствие возбуждения эта точка станет как бы отрицательным полюсом по отношению к невозбужденным частям, и тогда оба конца гальванометра окажутся связанными с электродами, лежащими на отрицательных и, следовательно, однородных точках мышцы, и ток покоя должен или исчезнуть, если потенциал места возбуждения по величине равен потенциалу у места повреждения, или он должен уменьшиться, если первый потенциал меньше второго. Здесь происходит борьба двух потенциалов: потенциала тока покоя и тока возбуждения. Этот последний ток по терминологии Германа — «ток действия», или «акционный ток», то есть ток, зависящий от состояния деятельности возбудимой ткани. В отношении электрофизиологических особенностей процесс возбуждения в представлении Германа приравнивается, таким образом, к процессу отмирания, ибо оба эти процесса превращают возбудимую ткань в источник электрических токов. Разница между этими процессами проявляется в том, что отмирание есть необратимый процесс, ведущий к смерти ткани, возбуждение же — обратимый процесс, за которым следует опять покой с возрождающейся способностью возбудимости. Приобрётшее популярность определение жизни, данное Клод Бернаром — «жизнь есть смерть» — получает на фоне электрофизиологических исследований своеобразное освещение в том отношении, что возбужденный участок ведет себя в деле образования электричества так, как отмирающий участок.
Дальнейшие успехи электрофизиологии задерживались из-за несовершенства гальванометров, которые если и были достаточно чувствительны, то все же имели недостаток в том отношении, что вследствие большой массы движущихся частей (магнитные стрелки в мультипликаторе, магнитный колокол в зеркальном гальванометре, катушка в магнитном поле гальванометра Д'Арсонваля, см. XII, 457) устанавливались слишком медленно на данную величину тока. При большом времени установки движение этих частей, недостаточно заглушенное, давало колебания собственного периода. Все это тормозило изучение электрических явлений для случаев одиночного мгновенного раздражения, ибо процесс возбуждения при одиночном импульсе раздражения протекает быстро и распространяется дальше по тканям с большой быстротой. Представим себе, что перед нами мышечный тяж, составленный из параллельных мышечных волокон. К двум неповрежденным точкам а и b (не дающим, следовательно, тока покоя) приложены электроды, отведенные к гальванометру (рис. 2). Если точку с раздражим одиночным индукционным электрическим током, то начавшееся здесь возбуждение побежит вдоль мышцы, и когда оно достигнет электрода а, то гальванометр d должен показать отклонение. Когда возбуждение, дальше, будет находиться между а и b, то гальванометр придет к нулю. При возбуждении точки b гальванометр даст отклонение в обратную сторону и при дальнейшем продвижении возбуждения придет опять к нулю.

Рис. 2.
Таким образом, гальванометр должен был бы дать движение, графическое изображение которого имело бы такой вид, как это изображено на рис. 2, то есть мы должны были бы получить так называемую двухфазную кривую с фазами 1 и 2, обращенными в разные стороны. Но перечисленные выше гальванометры не в состоянии указать токи от одиночного возбуждения, ибо вследствие быстротечности последних и при быстроте распространения их гальванометр с его инертными движущимися частями окажется совершенно беспомощным. Если бы мы раздражали не одним импульсом, а целым рядом их, то результат был бы такой же: гальванометр указывал бы покой. Допустим теперь, что один из электродов приложен к поперечному разрезу (рис. 3); в этом случае гальванометр даст вследствие тока покоя длительное отклонение от точки нуля.
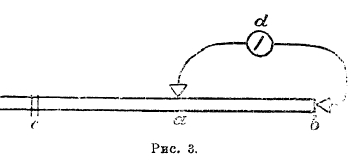
Рис. 3.
Одиночное раздражение точки с даст возбуждение, бегущее вдоль мышцы; достигнув точки а, возбуждение должно было бы дать кратковременное одиночное уменьшение тока покоя монофазного характера, но инертный гальванометр не может показать этого уменьшения; только в том случае, если раздражение будет быстро повторяться, точка а будет повторно становиться электроотрицательной, гальванометр покажет длительное уменьшение тока покоя, знакомое нам уже раньше, как отрицательное колебание. Но все же это будет извращенная передача, ибо на самом деле точка а испытывает не постоянный длительный процесс, а процесс колебательный, и гальванометр, если бы он был в состоянии следить за ходом изменений тока, должен был бы дать целый ряд колебаний. Вспомним, что подтверждением такого заключения может служить рассмотренный нами раньше вторичный тетанус.
Большим прогрессом в изучении указанных явлений было введение в электрофизиологию предложенного Бернштейном (Julius Bernstein, 1839-1917, профессор физиологии в Галле, написал: «Untersuchungen über den Erregungsvorgang im Nerven- und Muskel-System» (1871), «Lehrbuch der Physiologie» (1894) и др.) инструмента, так называемого дифференциального реотома. Этот инструмент позволял, правда, окольным, кропотливым путем, судить о всех изменениях электрических токов, сопровождающих одиночное раздражение возбудимой ткани, и даже позволял конструировать соответствующие кривые. Бернштейн сам, а также Герман выяснили при помощи этого инструмента, что ток действия возникает в раздраженной части уже в момент раздражения, то есть без скрытого периода, между тем как появление сокращения мышцы отодвинуто от момента раздражения на 0,01 секунды. По этой причине волна тока бежит впереди волны сокращения, причем скорость распространения обоих процессов одинакова — 3 метра в секунду. Удалось определить также для данной точки мышцы и длительность электрического процесса (0,03 сек.), а отсюда, принимая во внимание скорость распространения в 3 метра, можно было вычислить и длину электрической волны (3 Х 0,03 = 0,09 м). Для лягушечьего нерва было найдено, что длительность электрического процесса в каждой точке равна 0,002 секунды; принимая скорость распространения возбуждения по нерву 27 м в секунду, получаем для электрической волны 27 Х 0,002 = 0,054 м.
Период капиллярного электрометра и струнного гальванометра в электрофизиологии. Несмотря на серьезные результаты, достигнутые при помощи дифференциального реотома, прогресс электрофизиологии шел все же медленно, ибо инструмент этот имел много серьезных недостатков. Он совершенно оставлен теперь и имеет в настоящее время лишь историческое значение. Введение в физиологическую практику капиллярного электрометра, предложенного физиком Липманом (см. XLVIII, прил. современные деятели науки, 16), дало серьезный сдвиг в деле изучения животного электричества. Открылись совершенно новые, не предполагавшиеся раньше возможности. Капиллярный электрометр состоит из стеклянной трубки, конец которой оттянут в капиллярную трубочку. Налитая в такую трубочку ртуть не вытекает целиком: ртутный мениск в капилляре в состоянии нести на себе определенный столб ртути. Трубку погружают в сосуд, на дне которого налита ртуть, а сверху ртути — раствор серной кислоты. Таким образом, кислота находится между двумя ртутными массами, и в капилляре имеется граница между кислотой и ртутью. Если соединить какой-нибудь источник электровозбудительной силы с обеими ртутными массами, то в капилляре под влиянием поляризации, смотря по направлению тока, увеличивается или уменьшается кривизна ртутного мениска, а вследствие этого смещается в капилляре ртуть в одну или в другую сторону. Это смещение совершается с большой быстротой и не сопровождается собственными колебаниями. Смещение мениска может быть, конечно, наблюдаемо только при помощи микроскопа и может быть, что особенно важно, при помощи микроскопической проекции зарегистрировано фотографическим путем в виде кривой. Впервые французский физиолог Марей (Etienne Jules Marey, 1830-1904, профессор в Collège de France, изобретатель многочисленных саморегистрирующих приборов, применяемых в физиологии, написал: «La méthode graphique dans les sciences experimentales», 1878, «Physiologie du mouvement», 1890, «La chronophotographie», 1899, и др.) записал кривую тока лягушечьего сердца при помощи капиллярного электрометра; с большим совершенством делал позже такие записи английский физиолог Уоллер. В этих опытах электроды прикладывались непосредственно к обнаженному сердцу: ток действия, отведенный от неповрежденного сердца, давал двухфазную кривую; если один электрод касался поврежденного места, то получалась монофазная кривая. Без труда также удалось записать ток действия скелетной мышцы и нервных стволов при одиночном раздражении. Удалось записать ток действия от желез. Но все же многие явления еще не поддавались анализу, ибо установка капиллярного электрометра на данную величину потенциала совершалась, как оказалось при тщательных исследованиях, недостаточно быстро и чувствительность его была тоже недостаточно велика.
Совершенно исключительную роль в развитии современной электрофизиологии суждено было сыграть так называемому струнному гальванометру, построенному выдающимся исследователем животного электричества В. Эйнтговеном (см. XII, 458/59, и LI, 143). Этот инструмент дал возможность проникнуть значительно глубже в детали электрических явлений животного тела. В первый раз можно было записать ток от неповрежденных мышц животного и человека при одиночном раздражении. В первый раз можно было записать токи действия мышц человека при рефлекторном или волевом их сокращении. То, что много лет тому назад Введенский выслушивал при помощи телефона, то есть токи действия мышцы при волевом сокращении, можно было теперь получить в виде точной кривой. Число импульсов, высылаемых из центральной нервной системы, поскольку они обнаруживаются волевым тетанусом мышцы, можно было без труда в точности подсчитать. Электрофизиологические исследования нашли себе, благодаря струнному гальванометру, приложение в самых разнообразных областях физиологии. Вопросы об иннервации кожи и ее желез, пищеварительных желез, желез внутренней секреции, вопросы о функциях органов чувств получили на почве электрофизиологических изысканий новое освещение. Электрофизиология, стоявшая раньше в стороне от общего течения физиологической мысли и разрабатывавшая лишь теорию электрических токов животного тела с целью объяснения сущности процесса возбуждения, мало-помалу стала проникать, благодаря выработанной сильной методике, в самую глубь интересов общей и частной физиологии. Электрический ток, сопровождающий деятельное состояние всякой живой клетки, всякого органа тела, становится уже по одному этому сильнейшим орудием исследования функций организма. Мы являемся свидетелями того, как электрофизиологический метод перестает быть источником академического теоретизирования о сущности процесса возбуждения, каким он был во время Дюбуа-Реймона, и переходит на службу разработки насущных физиологических вопросов. Мало-помалу электрофизиологический метод проникает в клинику, именно в клинику нервных, мышечных и сердечных заболеваний.
Электрокардиография. А. Уоллер (Augustus Volney Waller, 1816-1870) впервые показал, что токи сердца можно отвести не только от обнаженного сердца, но и цельного животного и даже от человека. Он исходил из той мысли, что сердце в теле окружено тканями, пропитанными электролитами, проводящими электричество, и что сердечные токи выравниваются по этим проводникам. Если мы накладываем электроды, связанные с капиллярным электрометром, на тело животного или на его конечности, то ветви сердечных токов, проходящие по тканям, окружающим сердце, могут, хотя бы и в ослабленном виде, забежать в инструмент и проявить себя смещением ртутного мениска. Уоллеру, действительно, удалось впервые записать токи человеческого сердца. Запись эта была очень далека от совершенства, но в принципе, несомненно, вопрос был Уоллером решен. Впоследствии эти исследования перешли в руки Эйнтговена, который при помощи своего струнного гальванометра впервые получил точную, богатую деталями запись в виде кривой сердечных токов животного и человека. Эту запись он назвал электрокардиограммой (по предложению Самойлова она обозначается теперь сокращенно EKG). Исследования электрических токов сердца скоро оказались сильнейшим подспорьем при изучении физиологии и патологии сердца и дали начало самостоятельному отделу, так называемой электрокардиографии. Обозначение и форма EKG, по Эйнтговену, таковы: первый зубец Р (рис. 4) записывается током, сопровождающим сокращение предсердия; QRST соответствует токам при сокращении желудочка.
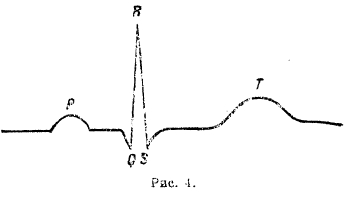
Рис. 4.
Уже одно то обстоятельство, что на EKG мы имеем отчетливое проявление деятельности и предсердий и желудочков, делает EKG ценной. Промежуток Р-R имеет большое значение: у нормального здорового человека он равняется 0,13-0,18 секунды. Отклонение за указанные пределы свидетельствует о болезненном состоянии сердца. Вся группа QRS длится нормально 0,06-0,08 секунды. Удлинение ее до 0,10 и больше уже свидетельствует о серьезных нарушениях функций сердечной мышцы. Нормальный вид EKG с правильно развитыми частями и зубцами Р на надлежащем месте, на надлежащем расстоянии от правильно очерченных QRS сразу исключает целый ряд заболеваний, подчас очень тяжелых. По указанию Эйнтховена EKG фотографируется в т. н. трех отведениях: I — от правой руки и левой руки, II - от правой руки и левой ноги и III — от левой руки и левой ноги. Таким образом, полученные EKG не одинаковы; между ними имеется совершенно определенная зависимость, указанная Эйнтховеном, а именно: если взять ординаты этих трех кривых для одного и того же момента, то ордината кривой II оказывается равной сумме ординат I и III кривой. Закон этот выражается так: II = I + III. Эйнтховен дальше показал, что на основании этих трех ординат можно для данного момента определить и т. н. электрическую ось сердца, т. е. равнодействующую направлений электрических потенциалов сердца для данного момента. Таким образом мы знаем, что в момент, когда зубец R дает свой максимум, RII = RI + RIII, и направление электрической оси идет справа и сверху, влево и вниз. Вместе с дыхательными движениями меняется вид для всех трех отведений, и можно показать, как при этом электрическая ось соответственно той или иной фазе дыхания меняет свое направление. Что касается происхождения EKG и ее отдельных частей, то выяснилось, что в общем зубцы QRS представляют выражение хода возбуждения по сердцу, а это в свою очередь связано с ходом т. н. проводящих путей сердца, т. е. узлов Флака, Тавара, пучка Гиса и т. н. ножек Тавара. Возбуждение начинается в синусовом узле Флака; возбуждение этого узла не улавливается струнным гальванометром. Отсюда возбуждение переходит на мышцы предсердия правого и левого. Как указано выше, прохождение по предсердию дает зубец Р, затем возбуждение переходит на узел Тавара, на пучок Гиса и на ножки Тавара; в течение всего времени хода возбуждения по указанным частям струна гальванометра находится на нулевой точке и в течение этого времени мышца предсердия к желудочков в покое и на кривой записывается прямая линия между Р и Q. Как только возбуждение пробегает от проводящей системы ножек к мышцам перегородки между желудочками, то сейчас же струна начинает писать желудочковый комплекс. Группа QRS свидетельствует о том, что возбуждение переходит с ножек на мышцы желудочков, и когда весь желудочек охвачен сокращением и все его точки одинаково возбуждены, то струна опять в покое и пишет нулевую линию между S и Т. После систолы начинается диастола, и тогда мышечные волокла постепенно переходят из состояния деятельности в состояние покоя:, в течение этого периода записывается зубец Т. Из сказанного нужно заключить, что вся часть QRST представляет собой комбинированную кривую из отдельных кривых правого и левого сердца. Винтерберг и Ротбергер первые делали на собаках опыты для выяснения вопроса об EKG правого и левого сердца. Они перерезывали либо правую, либо левую ножку Тавара. Если перерезать правую ножку, то возбуждение бежит с пучка Гиса по левой ножке, охватывает левый желудочек и потом бежит через мышечные волокна перегородки и вступает через 0,06 сек. в правый желудочек. Полученная при этом EKG (при втором и третьем отведении) имеет такой вид (см. рис. 5).
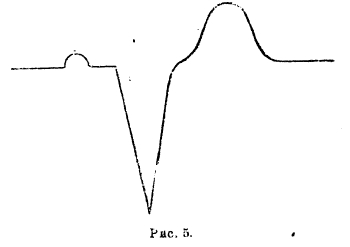
Рис. 5
Отклонение становится гораздо большим по величине, и кривая имеет строго дифазный вид с фазами, направленными в разные стороны. Вместо группы QRS, мы имеем один большой зубец, направленный книзу и длящийся дольше, чем нормальная группа QRS. Зубец Т, представляющий вторую, обратную фазу, не отделяется, как обычно, прямой линией от первой части кривой. При перерезке левой ножки указанные авторы получали кривую обратного вида (рис. 6).

Рис. 6.
Если алгебраически сложить обе кривые, левостороннюю и правостороннюю, сдвинув их правильно одну по отношению к другой, то получается группа QRS нормального вида, что и говорит в пользу того, что нормальная кривая представляет собой комбинацию двух кривых, право- и левосторонней. В патологических случаях у человека имеют место нарушения проводящих путей, например, в пучке Гиса, ножках Тавара; EKG дает возможность узнать, какое место проводящих путей пострадало. Кроме того, наблюдается целый ряд функциональных расстройств проведения возбуждения; все они распознаются с большой точностью по EKG. Точно так же могут быть установлены по EKG и места, в которых возникают так называемые экстрасистолы. Имеются формы болезней, которые, как, например, мелькание предсердий, были впервые поняты и выделены как таковые лишь на почве электрических кривых. Электрокардиография является, поэтому, одним из серьезнейших методов исследования сердечных заболеваний.
Современные теории электровозбудительных сил животного тела. Величина электровозбудительной силы для мышцы и нерва была уже установлена Дюбуа-Реймоном. Сравнительно большая величина в 20-30 милливольт для нерва и, в особенности, 40-80 милливольт для мышцы должна была представляться весьма загадочной, так как в животных цепях мы имеем дело лишь с проводниками второго рода; проводники первого рода — металлические части, без которых не обходятся гальванические элементы, не участвуют в электрической системе животного тела. Между тем мы знаем в животном теле не только указанные выше величины для электровозбудительной силы, но и гораздо большие: разность потенциалов в электрическом органе так называемых электрических рыб доходит до 100 вольт и больше, так что если соединить соответствующим образом тело рыбы с лампочкой накаливания, то в момент разряда электрического органа (сопротивление которого не больше 200 омов) лампочка вспыхивает. Как объяснить происхождение значительных электровозбудительных сил в теле животного? В настоящее время, благодаря успехам физики и химии, мы значительно приблизились к разрешению указанного вопроса: во всяком случае, теперь не трудно составить цепь из растворов электролитов, которая даст потенциал такого же порядка, какой дает и мышца. Верный путь в указанном вопросе электрофизиологии был найден с тех пор, как Нернст (см. XLVIII, прил. современные деятели науки, 17) впервые выяснил природу концентрационных и диффузионных цепей и дал для них уравнения. Для концентрационной цепи, состоящей из двух растворов соли, например, азотнокислого серебра, концентрации с1 и с2 и погруженных в них серебряных стержней, потенциал при температуре 18° равняется 58 log с1/с2 милливольт, если оба раствора соли соединены друг с другом так, что диффузионными токами можно пренебречь. Нетрудно усмотреть, что такая цепь с одинаковыми металлами дает, например, при с1: с2 = 10 напряжение, равное 0,058 вольт. Система такой концентрационной цепи в животном теле не может иметь места, так как металлических частей в теле нет. Ближе к условиям тела животного подходит система диффузионной цепи. Два раствора одного и того же электролита с концентрацией с и с1 дадут при соприкосновении потенциал, величина которого зависит от скорости, с какой ионы растворенного вещества будут устремляться из более концентрированного раствора в менее концентрированный. Для этого случая мы имеем формулы, определяющие величину потенциала:
(u-v)/(u+v)· 58 log (c1/c2)
милливольт, где u есть скорость катиона, а v — аниона. Для двух растворов азотнокислого серебра с отношением концентраций с1/с2 = 10 мы будем иметь не 58 милливольт, как при концентрационной цепи, а значительно меньше, ибо и для Ag — 54, а v для NO3 — 61,8, и поэтому величина и - v/и + v будет около 1 : 15 и, следовательно, потенциал нашей диффузионной цепи будет 58 · 1/15, то есть около 4 милливольт. Очевидно, что с увеличением разности скоростей обоих ионов будет расти диффузионный потенциал. Так как скорость водородного иона наибольшая, а именно 318, то ясно, что особенно большие потенциалы для диффузионной цепи будут в том случае, когда один из ионов есть водород, то есть в случае диффузионной цепи, составленной из двух растворов кислоты различной концентрации. Честь применения впервые этих данных физико-химии для цепей животного тела принадлежит русскому ученому В. Ю. Чаговцу (родился в 1873 г., профессор киевского университета, диссертация — «Очерк электрических явлений в живых тканях», в. 1-2, 1903-1906, и другие труды), который допускает, что в случае токов покоя, равно как и в случае токов действия, мы имеем систему кислоты о двух концентрациях. Уже Герман предполагал, что в альтерационном участке сосредоточена кислота. Чаговец исходил из предположения, что здесь нужно принимать участие угольной кислоты, скопляющейся в большой концентрации в измененном или в возбужденном участке. Идея Чаговца заключалась в том, что токи покоя или токи действия суть диффузионные токи. Так как известно, что раздраженная мышца производит в 6,5 раз больше угольной кислоты, чем покойная, то можно принять, что отношение концентрации угольной кислоты в обоих местах мышцы с1 : с2 равняется 6,5. Так как и для Н есть 318, v для НСO3 есть 40, то электровозбудительная сила будет
318 – 40/318 + 40 · 58 log 6,5,
то есть около 40 милливольт: получается потенциал такого же порядка, какой мы встречаем в мышце. Позднейшие авторы, однако, указывали, что во всей системе Чаговца есть допущения, с которыми трудно согласиться. На совершенно иную точку зрения стал Бернштейн, исходивший из предположения о существовании на поверхности возбудимых волокон перепонок со свойством полупроницаемости для ионов. Впервые эту идею высказал известный химик Оствальд (см. XLVIII, прил. современные деятели науки, 20). Он допускает существование в живом теле перепонок неодинаковой проницаемости для различных ионов и указывает, что такие перепонки могли бы объяснить происхождение больших потенциалов животного тела. Бернштейн отмечает, что мышца, как мы знаем, богата калийными солями, а жидкости тела и, в частности, кровь очень бедны этими солями. Поверхностная перепонка мышечных волокон, следовательно, не пропускает калийных солей.
Бернштейн видит причину этого в том, что из двух ионов калийной соли только катион проходит через перепонку, анион же пройти не может, и это обстоятельство ведет к появлению потенциала. Представим себе мышечный элемент в виде волокна (рис. 7).

Рис. 7.
В каждой точке образуется потенциал вследствие того, что катион с положительным знаком находится по одну сторону мембраны, а анион с отрицательным знаком по другую. Этот потенциал проявить себя, однако, не может, ибо вся система сама себя компенсирует в силу ее полкой симметрии. Если мы нанесем какое-нибудь повреждение в этой системе, тогда у поврежденного места разделения ионов больше не будет, система утратит свою симметрию, и получится ток с направлением от положительной неповрежденной поверхности к отрицательному поврежденному месту (рис. 8).
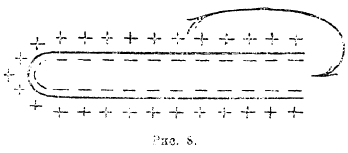
Рис. 8.
По теории Бернштейна всякое возбуждение изменяет проницаемость перепонки в сторону ее увеличения, как если бы здесь было вызвано повреждение: поэтому возбужденное место становится отрицательным по отношению к местам покойным. Следует обратить внимание на то, что схема Бернштейна предполагает предсуществование ионов по обе стороны перепонки и в этом отношении подходит больше к идеям Дюбуа-Реймона о «преэксистенции» электрических молекул, чем к идеям Германа об «альтерации», как причине потенциала. Бернштейн утверждает, что есть основание принимать в мышце скачек потенциала именно в местах неповрежденных на продольной поверхности, между тем как Герман принимает скачек потенциала в местах альтерации, то есть у поврежденного поперечника. Величину потенциала в мышце Бернштейн определяет, исходя из того, что в формуле диффузионного потенциала
![]()
милливольт он принимает скорость не пропускаемого иона за нуль; тогда величина потенциалов будет 58 log с1/с2 милливольт, то есть величина, характеризующая концентрационную цепь с металлами. Таким образом, мы видим, что при осуществлении некоторых условий можно ожидать для цепи, составленной из проводников второго порядка, таких же величин, какие дает концентрационная цепь с участием металла. На основании имеющихся анализов содержания калия в мышце и в крови Бернштейн приходит к вероятному потенциалу в мышце около 60 милливольт, что соответствует действительности. Совершенно новые точки зрения возникают на почве данных о так называемых фазовых потенциалах. Если мы растворим какой-нибудь электролит, например, НСl, в двух растворителях, не смешивающихся друг с другом, например, в воде и гваяколе, то электролит будет в зависимости от коэффициента разделения содержаться в одном и другом растворителе в различных концентрациях. На границе между обеими фазами возникает в таких случаях потенциал, который может достигать предельной величины, равной потенциалу концентрационной цепи, то есть 58 log с1/с2.
Если поместить органическое вещество, не смешивающееся с водой, между двумя растворами одной и той же соли концентрации с1 и с2, то на границе между фазами на одной и другой стороне получатся потенциалы, разность которых может достигать в пределе 58 log с1/с2. По совету знаменитого биолога Лёба (см. XLVIII, прил. современные деятели науки, 27) этим вопросом в применении к биоэлектричеству занялся Бейтнер, который построил ряд моделей, дающих потенциалы такой же величины, как животные возбудимые образования. В последнее время в лаборатории Гебера в Киле удалось получить системы фазовых потенциалов, где роль несмешивающегося с водой вещества между двумя электролитами играл слой белкового вещества, что значительно приближает эти модели к живым системам. Много интереса вызывают в последнее время модели, в которых местом появления потенциала служит перегородка между электролитами. Теорию этих потенциалов в мембране развил Доннан. Условия, при которых этот потенциал может появиться, — это непроницаемость перепонки для определенных ионов. Пусть по одну сторону перепонки находится раствор NaCl, ионы которого способны проникать через перепонку, а по другую сторону находится раствор натровой соли органической кислоты NaR, анион которой R не проходят через перепонку. Первоначальное состояние системы изменится вследствие диффузии, и наступит новое равновесие, при котором по одну сторону перепонки будут ионы Na, R, Cl, а по другую — ионы Na, Cl. Мы получим вследствие этого при установившемся равновесии большее количество ионов натрия в первой системе, чем во второй, а ионов хлора — в первой системе меньше, чем во второй. Эта разность концентрации ионов по одну и другую сторону мембраны вызывает разность потенциалов. Нужно думать, что такого рода потенциалы должны проявлять себя и в живых системах. В недавнее время Леман и Месман показали, что между кровью и водянистой влагой глаза у животного имеется разность потенциалов, которую следует понимать как потенциал Доннана, ибо кровь содержит белок, для которого сосуды непроницаемы, и потому не находится в водянистой влаге глаза, хлористый же натрий содержится в обеих жидкостях. Здесь даны, следовательно, все условия для образования потенциала Доннана. По анализу крови и влаги можно подсчитать количество ионов натрия и хлора в обеих жидкостях и вычислить потенциал. Прямое определение потенциала подтвердило теоретический подсчет.
Таким образом, можно с определенностью сказать, что если природа животных токов еще и не разгадана до конца, то, во всяком случае, современная физикохимия снимает с них покров их прежней загадочности и указывает верные пути для их детального исследования.
Благодаря новейшим методам осциллографии (Метьюз, Спиинс и др.) с усилителями оказалось возможным регистрировать токи действия в отдельных волокнах и установить значительное различие в возбудимости, скорости проведения возбуждения и в амплитуде тока действия. Это различие стоит в связи с толщиной нервного волокна: чем последнее толще, тем интенсивность вышеуказанных процессов выше, и наоборот. На первом месте стоят двигательные (эфферентные) волокна, чувствующие (афферентные) уступают им и в свою очередь дают известные градации. Удалось, далее, показать, что ток действия протекает в фазах различной силы значительно дольше (до 5 секунд), чем считалось раньше, когда могли констатировать лишь первую, наиболее сильную фазу, длящуюся всего 0,001-0,002 секунды. Наконец, современные методы дают возможность регистрировать токи действия коры головного мозга, то есть получать так называемую электроцереброграмму.
Литература: Е. Du-Bois-Reymond, «Untersuсhungen über tierische Elektrizitüt», Berlin, 1848-49, и «Gesammelte Abbandlungen zur allgemeinen Muskel- und Nerven physik», Leipzig, 1875; Сеченов, И. М., «Лекции о животном электричестве», СПБ, 1862; Hermann, L., «Hаndbuсh der Physiologie». Bd. I, Th. 1, Leipzig, 1879 (есть русский перевод); Biedermann, W., «Electrophysiologie», Vena, 1895, и «Ergebnisse d. Physiologie», Bd. I. Th. 2, 1902, и Bd. II, Th. 2, 1903; Bose, I. Ch., «Comparative Elektrophysiologie», London, 1907; Cremer, М., «Nagel's Handbuch der Physiologie», Bd. IV, Braunschweig, 1909; Bernstein, I., «Elektrobiоlogie», Braunschweig, 1912; Garsten, S., «Wiuterstein's Handbuch d. vergleich. Physiologie», Bd. III, 1910-14; Bewner, R., «Die Entstehurg elektr. Ströme im lebenden Gewebe», Stuttgart, 1920, и «Bioсhemische Zeitchr.», Bd. 137. 1923; Лазapeв, П. П., «Ионная теория возбуждения», Госизд., 1923; Höber, R., «Zeitschr. f. physikal. Chemie», 110, 1924. Статьи Steinnausen, W., Cremer, M. и Rosenberg, Н., в «Bethe’s Handb. d. normalen und pathel. Physiologie», Вd. 8, 2 Häfte, Berlin, 1928; Schäfer, Н., «Neuere Untersuchungen über den Nervenaktionsstrom. Ergebisse d. Physiologie», Bd. 36, 1934.
А. Самойлов.
| Номер тома | 53 |
| Номер (-а) страницы | 305 |