Эмбриология
Эмбриология (греческий έμβρυον — зародыш, λόγος — учение), наука о развитии особи, об индивидуальном развитии организма после того, как он обособился от материнского. При половом размножении отправным пунктом развития является яйцеклетка, при бесполом или вегетативном — различные многоклеточные зачатки (почки кишечнополостных, мшанок), группы недифференцированных, эмбриональных клеток (интерстициальные клетки хобота гидромедуз, археоциты в геммулах губок, статобласты мшанок) и даже одиночные эмбриональные клетки (необласты в зонах деления малощетинковых кольчатых червей и т. п.). В последнем случае кардинальное отличие от яйцеклетки заключается в отсутствии редукционного деления, которое при половом размножении всегда налицо, хотя бы в рудиментарной форме (при партеногенезе). Завершением развития особи считают обычно наступление половой зрелости, но если не говорить даже о таких исключительных случаях, как диссогония гребневиков, у которых половая зрелость наступает дважды, или как довольно распространенный педогенез (размножение в молодом возрасте, в личиночном, например, состоянии), то и тогда придется признать, что с наступлением половой зрелости развитие не заканчивается; некоторые членистоногие продолжают, например, линять после спаривания. Быть может, было бы правильнее под развитием особи понимать весь период жизни, до естественной смерти; но практически эмбриолога занимает обычно период наиболее интенсивного формирования (морфогенеза), приходящийся на развитие «в яйце» и нередко сверх того на «постэмбриональное» развитие, если последнее сопровождается глубокими изменениями («метаморфозом» личинки).
Эмбриологию нельзя целиком отнести к области морфологии или к области физиологии, так как наряду с морфологическим подходом к изучению развития особи (описательная, сравнительная, филогенетическая эмбриология) существует и физиологический, выясняющий экспериментальными методами физиологические причины индивидуального развития (механика или физиология развития, экспериментальная морфология). Из других биологических дисциплин эмбриология имеет тесное соприкосновение с филогенетикой, с генетикой и с учением об эволюции. Связь с филогенетикой, изучающей пути эволюционного развития организмов, восстанавливающей их родословную, осуществляется в силу того, что по ходу зародышевого развития можно до известной степени судить об истории происхождения данного организма: зародышевое развитие является как бы историческим документом для филогенетики. Связь с учением о наследственности, генетикой, осуществляется через посредство феногенетики, изучающей физиологию развития тех признаков, изучение закономерностей наследования которых составляет задачу генетики. Наконец, изучение соотношения между индивидуальным развитием и развитием историческим, эволюционным имеет не только значение подсобное для филогенетических построений, как это всеми, к сожалению, считается, но и гораздо более широкое и глубокое значение в смысле выяснения закономерностей эволюции вообще.
История эмбриологии. Из авторов древнего мира наибольший интерес представляет Аристотель (384-322 гг. до нашей эры); его разработка вопросов размножения животных имела важное значение для развития эмбриологии. Аристотель различал три способа возникновения особей: самопроизвольное зарождение, бесполое размножение и половое. При самопроизвольном зарождении одухотворенная природа создает низших животных из гниющих веществ, например, комаров и стрекоз из ила; при бесполом размножении новые особи возникают путем почкования. Половое размножение сопровождается участием двух особей — мужской и женской; последняя, менее совершенная, дает лишь материал для построения зародыша; ее «семя» (менструальная кровь) не достигает той ступени развития, как семя мужской особи; последнее одухотворяет материю, предоставляемую женской особью, дает ей движение и форму; часть семени остается неизрасходованной на формирование зародыша и сохраняется у зародыша, чтобы впоследствии служить для размножения по достижении половой зрелости. Наиболее совершенно развитие прямое, свойственное млекопитающим; акулы, образующие яйца, развивающиеся в теле матери, стоят уже ниже; откладывающие яйца птицы и пресмыкающиеся занимают на лестнице еще более низкое положение; далее идут костистые рыбы, лягушки, головоногие моллюски, яйца которых не столь совершенны, как у предыдущих животных, и развитие продолжается по выходе из яйца и, наконец, ниже всего стоят насекомые, которые откладывают даже не яйца, а личинок, лишь потом превращающихся в яйца (окукление).
Огромный период от Аристотеля и кончая средними веками, мало дал для развития науки вследствие религиозной нетерпимости, препятствовавшей не только дальнейшему изучению природы, но и использованию наследия дохристианских авторов.
В эпоху Возрождения вопросы размножения животных живо интересовали исследователей, но наибольшее развитие проблема получила только у Гарвея (1578-1657; см.), которому принадлежит знаменитое изречение «все из яйца». Гарвей старался отыскать начало, отправной пункт индивидуального развития животных; при этом он не ограничивался изучением авторитетов древности, подобно весьма многим ученым своей эпохи, но обратился к самостоятельным обширным исследованиям развития крупного яйца и зародыша млекопитающего (косули). В результате, в молодом зародыше млекопитающего, развивающемся в матке, он признал образование, соответствующее яйцу птиц. Развитие высших животных сопровождается дифференцировкой первоначально однородной материи (эпигенез), у низших же животных развитие заключается лишь в развертывании готовых зачатков (выход насекомого из куколки, которую принимали за яйцо). Зарождение самопроизвольное, не из яйца, Гарвей все-таки допускал для самых низших животных. Некоторые идеи Гарвея были разработаны Граафом (1641-1673), исследовавшим строение половых органов. В то время как относительно яичников птиц давно уже было известно, что в них образуются яйца, яичники млекопитающих считались семенниками, а относительно «женского семени» существовали противоречивые представления. Грааф обосновал взгляд своего учителя Ван-дер-Горна, что фолликулы (Граафовы пузырьки) суть яйца, назвал «женский семенник» яичником, а открытые Фаллопием протоки (Фаллопиевы трубы) признал за яйцеводы, констатировав, что по ним яйца проходят в матку.
Что касается спермы, то весьма долгое время ученые считали совершенно невероятным, чтобы ограниченное количество жидкой материи могло из влагалища или клоаки проникнуть до мест нахождения молодых яиц и оплодотворить последние. Поэтому большим распространением пользовалось исходившее от Аристотеля представление о том, что сперма, как материя, в оплодотворении не играет роли, но она порождает некое духовное начало, проникающее в яйцо, хотя бы последнее находилось далеко и было покрыто оболочками. Лишь в 1677 году применение шлифованных линз для изучения невидимых глазу объектов привело Левенгука и Гамма к открытию в семенной жидкости спермиев, обладающих активным движением, но это открытие еще далеко не означало, что роль спермиев в оплодотворении получила общее признание.
Наблюдения первых микроскопистов (Мальпиги, 1628-1694, Сваммердам, 1637-1680) над строением личинок и куколок насекомых, над строением зародышей позвоночных (цыпленка в яйце) обнаружили неожиданно большую сложность этих объектов. Подобная же, но лишь невидимая сложность была приписана и недавно открытым спермиям: раз они двигаются, они должны обладать мускулатурой, нервной системой и т. п.
Обсуждение накопившихся наблюдений в течение XVIII века привело, в связи с влияниями со стороны некоторых философских учений, к концепции преформизма: в материальном зачатке, из которого развивается особь, все предобразовано, и развитие заключается лишь в развертывании и росте существующих уже частей. Завершением этой точки зрения было представление о вложении в зародыша зародышей всех будущих поколений; некоторые объекты первых микроскопистов (например, Volvox), казалось, давали фактическое подтверждение этой гипотезе. Но где находится переформированный зародыш: в яйце или в мужском семени? Большинство (овисты: Грааф, Мальпиги, Валлиснери и др.) склонялось к первому предположению, и спермии рассматривались в таком случае как случайные обитатели семенной жидкости, как паразиты; с точки зрения других (анималькулистов: Левенгук, Гартсекер, Андри и др.), зародыш заключен в сперматозоиде, а яйцо служит лишь питательной средой. Открытие партеногенеза у тлей (Бонне) поддержало позицию овистов. Лишь в конце XVIII века опытами Спаланцани с фильтрацией спермы было положено начало взгляду о материальном значении спермы, ее сперматозоидов в процессе оплодотворения.
Против теории преформизма выступил в 1759 году Каспар Фридрих Вольф (см.) в своей работе «Theoria generations»; основываясь на наблюдениях над развитием цыпленка в яйце, Вольф утверждал, что развитие сопровождается новообразованиями, что сложный дифференцированный организм формируется из однородного зачатка под влиянием питания и роста, что до начала развития яйцо вообще даже не имеет никакой структуры, являясь однородной массой. Эта эпигенетическая точка зрения, доведенная у Вольфа до крайности, не имела успеха у современников, но в истории эмбриологии сыграла свою роль (ср. анатомия, II, 601/02).
Среди естествоиспытателей ХVІІ, XVIII и начала XIX веков широко было распространено учение о «серии» или «лестнице» организмов, о том, что все растения и животные располагаются по степени сложности и совершенства в один непрерывный ряд. Это учение, распространенное в разные времена и во многих странах, в указанный выше период особенно развилось под влиянием идеи о непрерывности всего сущего, развитой в философии Лейбницем (1646 -1716) и Робине (1735-1820). Наиболее ярким представлением теории лестницы организмов в XVIII веке является Бонне (1720-1793), у которого самые нижние ступени лестницы занимают огонь, воздух, вода и земля, а самые высокие — четвероногие, человек, чины небесной иерархии и, наконец, бог. Возникшее в последнем десятилетии XVIII века и широко распространенное в первой четверти XIX века натурфилософское направление в биологии восприняло идею лестницы и распространило ее на зародышевое развитие: зародыш высшего животного последовательно проходит в своем развитии через состояния, соответствующие взрослым состояниям современных низших животных, стоящих ниже его на лестнице (теория параллелизма; Меккель, Окен, Серр, Ратке и многие другие). Если в эмбриологии идея непрерывности нашла свое выражение в теории параллелизма, то в сравнительной анатомии она вела к теории единого плана строения у всех животных (Э. Жоффруа Сент-Илер), а в систематике — к отрицанию реальности не только систематических групп высшего порядка, но и видов: нет ничего, кроме непрерывной цепи особей. Развитие науки привело к крушению всех этих идей, к утверждению постоянных видов (Линней, 1735), постоянных типов (Кювье, 1812), господствовавшему до Дарвина.
В области эмбриологии с теорией параллелизма было покончено главным образом благодаря классической работе Бэра (см.), вышедшей в 1828 году. Бэр явился обоснователем теории типов в эмбриологии, так же как Кювье — в сравнительной анатомии; тот и другой установили одни и те же 4 типа: позвоночных, членистых, моллюсков и лучистых. Каждому из этих типов соответствует, по Бэру, и особый тип зародышевого развития; у позвоночных, прежде всего, развивается спинная сторона тела с зачатком нервной системы в виде борозды; у членистых, наоборот, брюшная сторона с находящейся на ней нервной системой формируется раньше, а спинная сторона замыкается позже; у моллюсков рано появляется зачаток раковины; у лучистых — лучевая симметрия тела. Далее, Бэр пришел к выводу, что принадлежность животного к определенному типу выявляется у зародыша очень рано; вследствие этого зародыш позвоночного вовсе не проходит в своем развитии через низшие типы. Вообще никакого прохождения через взрослые состояния низших животных нет, но имеет место последовательное обособление зародыша от стадии, свойственной всему данному типу, к стадии, типичной для класса, потом для отряда и т. д. Зародыш человека сперва является только зародышем позвоночного, потом зародышем, имеющим аллантоис, потом зародышем млекопитающего и т.д., последовательно обособляясь сперва от остальных трех типов животных, потом от жаберных позвоночных, потом от Sauropsida и т. д. Расхождение зародышей возрастает, таким образом, с ходом развития. Зародыш может быть похож только на зародыш другой группы, но никогда не на взрослое животное. Сходство между зародышами больше, чем между взрослыми формами, и тем больше, чем более ранние стадии сравниваются.
Широкое распространение явлений зародышевого сходства было выдвинуто Дарвином в «Происхождении видов» (1859) в качестве доказательства эволюции. Дарвин нашел «закону зародышевого сходства» естественное объяснение с точки зрения эволюционной теории: эволюция связана с изменениями в ходе зародышевого развития; эти изменения впервые появляются обычно не на самых ранних стадиях: ранние стадии остаются, таким образом, незатронутыми и сохраняют сходство с развитием предков.
После появления «Происхождения видов» исследователи стали обращаться к изучению родственных отношений между группами, к выяснению их родословных, путей их эволюции, с целью укрепления эволюционной идеи в период борьбы за дарвинизм и дальнейшей разработки эволюционной теории. Пионером этого филогенетического направления, широко распространившегося в семидесятых и восьмидесятых годах, явился Фриц Мюллер (1821-1897, один из первых сторонников Дарвина, в 1864 году опубликовал «Für Darwin», где обратил внимание на филогенетический закон), занявшийся филогенией в группе ракообразных, но пришедший к гораздо более широким и важным выводам, относящимся к вопросу о соотношении индивидуального развития и эволюции. Фриц Мюллер, принимая вместе с Дарвином, что эволюционные изменения индивидуального развития впервые проявляются не на самых ранних стадиях, пришел к выводу, что эти изменения могут наступить либо до достижения животным состояния взрослых родителей, либо непосредственно после; во втором случае в конце развития прибавляется новая стадия, и вследствие этого в развитии потомков проходятся стадии, бывшие у предков достоянием взрослого организма (1864).
Значение изучения зародышевого развития для понимания истории происхождения данного животного особенно пропагандировалось Геккелем (см.) в 1866 и в 1874 годах, выдвинувшим тезис о повторении «онтогенией» «филогении» и возведшим это положение в «основной биогенетический закон». Задавшись целью выяснить родословную человека путем изучения его зародышевого развития, Геккель очень скоро натолкнулся на такие черты зародышевого развития, которые никоим образом не могли считаться свидетельствами об историческом прошлом, не могли быть приписаны предкам. В виду этого, Геккель пришел к совершенно правильной мысли о том, что в развитии особи имеются черты палингенетические (см. ценогенетические признаки), имеющие значение исторических документов, и черты ценогенетические, искажающие, стирающие историческую верность онтогенеза. Однако, отсутствие точных и продуманных формулировок привело к тому, что вопрос о том, какие признаки в каждом конкретном случае считать палингенетическими и какие ценогенетическими, сделался объектом произвола исследователя. Тезис Геккеля о том, что филогенез является «механической причиной» онтогенеза, также имел немало дурных последствий; стали представлять себе, что новообразования в эволюции появляются сперва во взрослом состоянии, а потом сдвигаются на все более ранние стадии зародышевого развития, изменяя ход последнего. После Геккеля теория рекапитуляции стала привлекать к себе большое внимание исследователей, причем постепенно выявились два основных учения: одни исследователи возвращаются назад, к Бэру, скептически относятся к возможности надставок новых стадий и к рекапитуляции признаков взрослых предков (О. Гертвиг, Нэф и др.), другие, наоборот, разрабатывают дальше положения Фрица Мюллера и Геккеля (Вейсманн, Северцов, Франц и др.).
Особенно основательной переработке эти положения подверглись со стороны А. Н. Северцова, который на основании обширных исследований по развитию позвоночных в ряде работ (начиная с 1910 г.) обосновал учение о филэмбриогенезах. По Северцову, эволюционные изменения хода индивидуального развития впервые появляются не во взрослом состоянии, когда развитие в основном уже закончено, а именно во время самого развития, в период энергичного формирования, что и ведет к изменению признаков взрослых животных; для того, чтобы изменился признак взрослого животного, нужно, чтобы изменился ход развития этого признака. Какие-либо «сдвигания» признаков от взрослого состояния в зародышевое развитие и не доказаны, и не нужны. Не все то, что ново, что недавно изменило ход зародышевого развития, является ценогенезом; ценогенезы суть такие изменения зародыша или личинки, которые на взрослом состоянии не отражаются, для эволюции взрослых форм значения не имеют. Если же изменение зародыша ведет к изменению взрослого животного, то это не ценогенез, а филэмбриогенез, то есть филогенетическое изменение зародышевого развития, на котором строится эволюция.
Влияние филогенетического направления на развитие эмбриологии было весьма значительно; сравнительная эмбриология особенно продвинулась вперед благодаря работам русских исследователей: И. И. Мечникова (1845-1915) и А. О. Ковалевского (1840-1901). Много внимания было уделено эмбриологии беспозвоночных, и представление Бэра о том, что самые первые стадии развития у всех животных одинаковы, а за ними следуют стадии, совершенно несравнимые в различных типах, было оставлено. Прогрессивное для своего времени, периода борьбы с теорией параллелизма, учение Бэра о постоянстве четырех типов было отвергнуто с победой эволюционной теории. Возросший в связи с филогенетическим направлением интерес к эмбриологическим исследованиям привел к накоплению огромного фактического материала в различных областях эмбриологии.
Первые стадии развития. При половом размножении многоклеточных животных развитие особи начинается с деления, или, как говорят обычно, с дробления яйца; это дробление отличается от обыкновенного клеточного деления тем, что оно не связано с ростом получающихся в результате дробления клеток, так называемых бластомеров, и последние с каждым новым делением становятся все более мелкими. Яйцеклетка содержит обычно запас питательных веществ, идущих на питание развивающегося зародыша и называемых желтком; количество и в особенности расположение желтка в яйцеклетке оказывает значительное влияние на ход дробления, так как желток представляет собой балласт, задерживающий дробление тех частей яйца, в которых он сосредоточен (ср. животные, XX, 233/34).
Различают яйца гомолецитальные — с не очень большим количеством равномерно распределенного по плазме желтка (многощетинковые, иглокожие, млекопитающие); яйца телолецитальные – с большим количеством желтка, оттесняющего активную, образовательную плазму к одному полюсу – анимальному, противоположному богатому желтком вегетативному полюсу (рыбы, амфибии, рептилии, птицы); свойственные большинству насекомых центролецитальные яйца имеют скопление образовательной плазмы в центре (где лежит ядро) и по самой периферии, желток же помещается в промежуточных областях плазмы.
Когда содержание желтка в яйце достигает известных пределов, яйцо уже не может дробиться полностью и части его, особенно богатые желтком, не разделяются или запаздывают с разделением на бластомеры; такое дробление называется частичным. Частичное дробление называется дискоидальным, если яйца телолецитальны, и дроблению подвергается в первую очередь лишь анимальный полюс, покрывающий дисковидной шапочкой остальную богатую желтком массу яйца (рис. 1);
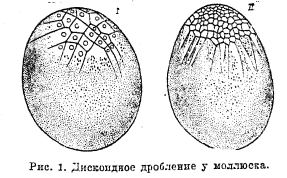
Рис. 1. Дискоидное дробление у моллюска.
оно называется поверхностным у центролецитальных яиц, у которых ядро, находящееся первоначально в центре, делится несколько раз, дочерние ядра выходят в поверхностный слой плазмы, и в последнем разыгрываются ближайшие последующие этапы развития (рис. 2).
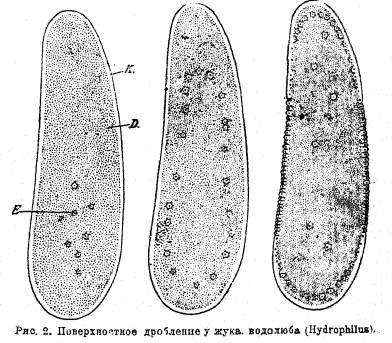
Рис. 2. Поверхностное дробление у жука водолюба (Hydrophilus).
Полное дробление наблюдается у гомолецитальных яиц и у телолецитальных; в последнем случае оно является неравномерным, так как бластомеры получаются неодинаковой величины: более быстро делящиеся бластомеры анимального полюса получаются мельче (микромеры), чем бластомеры вегетативного полюса (макромеры, см. рис. 3). Полное равномерное дробление см. животные, XX, табл. 5, рис. 3.
Кроме классификации явлений дробления в зависимости от желтка, различают еще типы дробления по признаку направления оси веретена деления ядра, по расположению бластомеров друг относительно друга. При радиальном, или лучевом, типе дробления вследствие деления яйца по меридианальным плоскостям, проходящим через вертикальную ось яйцеклетки, бластомеры располагаются в радиальных направлениях вокруг этой оси (рис. 4). Двусторонний тип дробления характеризуется тем, что яйцо разделяется плоскостью дробления на две симметричные половины, в каждой из которых дальнейшие этапы дробления протекают одинаково. При спиральном типе дробления бластомеры располагаются в радиальных направлениях вокруг оси, но в вертикальном направлении они лежат при этом не друг над другом, а чередуясь (рис. 5).
В результате дробления чаще всего образуется зародыш, представляющий собою округлую группу клеток, располагающихся вокруг центральной полости в один слой; эта стадия называется бластулой, а полость – бластоцелем (см. животные, ХХ, табл. 5, рис. 4). Реже получается не бластула, а морула, отличающаяся отсутствием полости и расположения бластомеров в один слой.
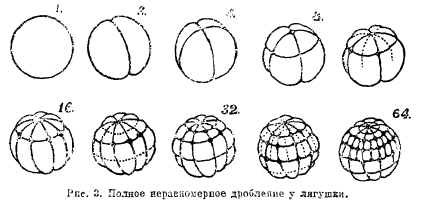
Рис. 3. Полное неравномерное дробление у лягушки.
Бластоцель может быть также не выражен вследствие впячения в него крупных макромеров вегетативного полюса или заполнения его нераздробившимся желтком (ср. животные, XX, 234/36).
Бластула является однослойным зародышем; в дальнейшем зародыш переходит в двуслойную стадию — гаструлу (см. животные, XX, табл. 5, рис. 5). Гаструляция осуществляется довольно разнообразными способами. Способ инвагинации заключается в том, что на вегетативном полюсе стенка бластулы впячивается внутрь; этот впяченный внутренний слой называется энтодермой и окружает первичную пищеварительную полость, открывающуюся наружу отверстием, называемым гастропором, или бластопором. Слой, одевающий зародыш снаружи, называется эктодермой; такие клеточные зародышевые слои вообще носят название зародышевых листов, или пластов. В тех случаях, когда бластула состоит из макро- и микромеров и лишена полости, инвагинация невозможна, и двухслойность получается в результате обрастания эпиболии быстрее делящимися микромерами инертных макромеров; получается зародыш с энтодермой из макромеров внутри и эктодермой из микромеров снаружи, но без гастроцеля.
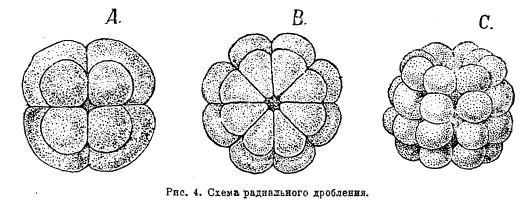
Рис. 4. Схема радиального дробления.
Деляминацией называется такой способ гаструляции, при котором клетки бластулы, делясь, отделяют от себя дочерние клетки внутрь бластоцеля; эти отделившиеся внутрь клетки и образуют энтодерму.
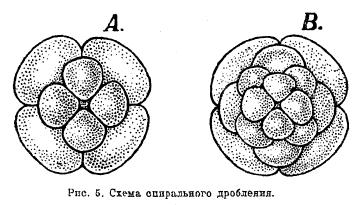
Рис. 5. Схема спирального дробления.
При иммиграции часть клеток стенки бластулы покидает свои места и погружается в бластоцель, образуя там энтодерму. Нередки переходы и комбинации между различными способами гаструляции.
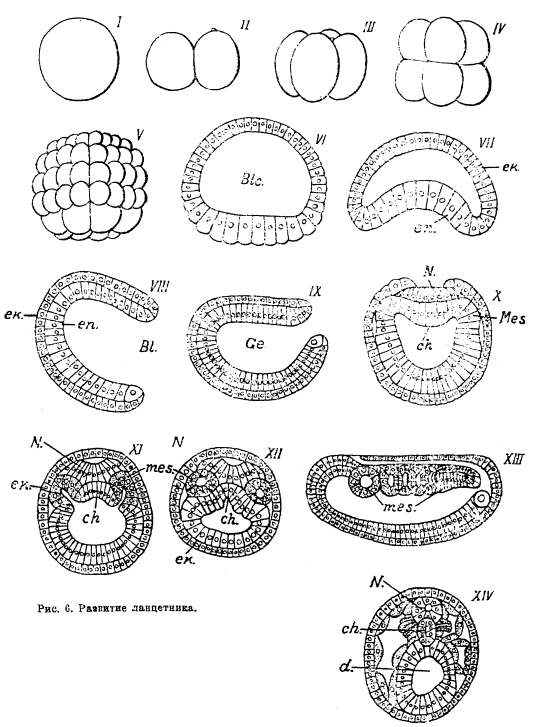
Рис. 6. Развитие ланцетника.
Из эктодермы развивается однослойный кожный эпителий у большинства животных и многослойный эпидермис у высших хордальных; различные кожные железы, кутикулярные образования, панцири, раковины, перья, волосы, когти, копыта и т. п., будучи производными кожи, также развиваются преимущественно из эктодермы. Из эктодермы же развиваются нервная система и органы чувств, по крайней мере, наиболее существенные — воспринимающие части последних; так, например, у хордальных центральная нервная система закладывается в виде кожного желобка вдоль спинной стороны тела зародыша, потом этот желобок погружается и замыкается в трубку, полость которой сохраняется во взрослом состоянии в форме спинномозгового канала и желудочков головного мозга (рис. 6, N). Сетчатка глаза хордальных возникает как выпячивание стенки головного мозга, внутреннее ухо обособляется от кожи в виде слухового пузырька. Энтодерма образует внутреннюю выстилку кишечного канала, большей частью на значительном его протяжении; так, у хордальных лишь небольшая часть ротовой полости и задней (прямой) кишки выстланы эктодермой, заворачивающейся снаружи, от кожного покрова. Различные придаточные полости и органы, возникающие у зародыша как выросты энтодермальной части кишечника, также, конечно, являются производными энтодермы; у хордальных такими производными являются легкие и пищеварительные железы.
Большинство животных трехслойно и, кроме экто- и энтодермы, имеет еще средний зародышевый слой, или мезодерму, возникающую обычно в той или иной связи с энтодермой. У хордальных мезодерма образуется в виде мешковидных выростов первичного кишечника, симметрично расположенных справа и слева от него (рис. 6, Х-XII). Эти выросты отшнуровываются, превращаясь в полые пузыри, так называемые сомиты. Сомиты правой и левой стороны, разрастаясь и встречаясь друг с другом в плоскости симметрии тела (сагиттальной), заполняют все пространство между кожей и зачатком кишечного канала; их полости превращаются во вторичную полость тела или целом, в котором лежат внутренние органы; их наружные стенки, примыкающие изнутри к эктодерме (соматический листок), дают соединительнотканный слой кожи (cutis, или дерма); их внутренние стенки (висцеральный листок), прикладывающиеся снаружи к зачатку кишечника, образуют соединительнотканную и мышечную обкладки кишки, а также мезентерий, на котором кишечник подвешен к стенке тела; выстилающий полость тела перитонеальный эпителий также происходит из мезодермы (рис. 6, ХIV). Но этим роль мезодермы в эмбриогенезе, особенно хордальных, далеко не ограничивается; из мезодермы возникает мускулатура, кровеносная система, мочеполовая система, скелет. У низших хордальных скелет представлен хордою, плотной осью клеточного строения, которая тянется вдоль тела под нервной трубкой и над кишечником и в эмбриональном развитии отделяется от первичного кишечника (рис. 6, Х-XIV); у высших хордальных (позвоночных) хорда имеется у зародыша, но на более поздних стадиях заменяется, вытесняется позвонками, хрящевыми или костными, мезодермического происхождения. Из других типов животных мезодермический скелет имеют иглокожие. О развитии человеческого зародыша см. зародыш, XX, 612/19.
Механика развития. С конца XIX века в эмбриологии начинает получать широкое распространение экспериментальное направление, известное под различными наименованиями: «механика развития», «экспериментальная морфология», «экспериментальная эмбриология», «физиология развития»; из этих названий наиболее распространенным и укрепившимся является первое. Основоположником этого направления считается германский анатом Вильгельм Ру (1850-1924) основавший журнал «Архив механики развития организмов» в 1894 году. Задачей механики развития является выяснение физиологических причин развития особи путем экспериментального анализа.
Достигнутые результаты побуждают смотреть на организм, как на расчлененное целое, части которого находятся в связи и взаимодействии, как на сложную систему, развивающуюся во взаимодействии с окружающей средой; старый спор об эпигенезе или преформизме в развитии особи в значительной мере снимается в свете новых данных.
Разнообразные опыты с дробящимися яйцеклетками, произведенные Ру, Гертвигами, Дришем, Бовери, Морганом и другими (разделение первых бластомеров, изоляция отдельных бластомеров и их групп, изменение расположения бластомеров, перетяжка яйца, умерщвление части бластомеров и т. п.), показали, что яйцеклетка является отнюдь не массой однородного вещества, но обладает такой структурой плазмы, которая обусловливает первые этапы развития. Эта структура не заложена в яйцеклетке с самого начала в готовом виде, но сама развивается в яйце; дробление застает эту структуру у разных животных то более, то менее сформированной. Поэтому у одних животных один из первых двух или четырех бластомеров, будучи изолирован, развивается как целое яйцо, то есть дает нормальный зародыш; у других животных структура плазмы яйцеклетки складывается раньше, и изолированный бластомер дает лишь то, что он дал бы, находясь в составе целого: половинный, четвертной зародыш или, по крайней мере, половинное развитие некоторых органов. В первом случае яйца называются регуляционными и встречаются у гидромедуз из кишечнополостных и у вторичноротых (иглокожие, амфибии), трехслойных животных; во втором случае яйца называются мозаичными и свойственны гребневикам из кишечнополостных, а из трехслойных — первичноротым (кольчатые черви, моллюски). Оба типа яиц и дробления не резко разграничены и возможны переходы; второй тип дробления называют также детерминированным, так как к моменту изоляции бластомеров путь их развития уже оказывается определен, детерминирован.
Изучение дальнейших стадий зародышевого развития, закладки систем органов и отдельных органов и их частей экспериментальными методами (главным образом методом пересадки, трансплантации зачатков у амфибий) показало, что развитие одних частей зародыша происходит при участии влияний со стороны других частей, что самодифференцировка зачатка в то же время является до известной степени зависимой дифференцировкой. Так, например, по данным Шпемана, Мангольда, Бауцмана и др., зачаток центральной нервной системы у амфибий развивается, испытывая какое-то стимулирующее влияние со стороны подстилающей его снизу мезо-энтодермы, дающей впоследствии хорду, мезодерму и спинную стенку кишечника; если же проследить, откуда берет начало эта стимулирующая часть зародыша, то оказывается, что таким местом является зона, примыкающая к бластопору со спинной стороны; она является организатором или организационным центром по отношению к зачатку центральной нервной системы и, будучи пересажена в необычное место зародыша, может вызвать образование нервной системы в эктодерме данного участка, может детерминировать эктодерму, направив ее развитие по пути образования нервной системы. Нервная система дифференцируется на отделы; так называемый промежуточный мозговой пузырь образует два боковых пузыреобразных выпячивания – зачатки глаз; глазные пузыри путем вдавления обращенной кнаружи стенки превращаются в двуслойные бокалы, и, когда бокал вплотную подходит к коже зародыша, он стимулирует образование хрусталика, последний же после своего отделения от кожи индуцирует превращение кожи в роговицу. Таким образом, намечается целая цепь процессов развития, все более и более специальных, причем опережающие в развитии части определяют направление развития некоторых более поздно развивающихся зачатков, детерминируют, организуют те зачатки, развитие которых должно быть с ними согласовано. Зависимая дифференцировка не ограничивается зародышевым периодом развития, но и в течение постэмбрионального развития моменты энергичного формообразования сопровождаются стимуляцией одних частей тела со стороны других. В настоящее время целый ряд тем из огромной области индивидуального развития уже успел подвергнуться основательной разработке с точки зрения механики развития. Упомянем выяснение причин клеточного деления; открыто особое излучение, так называемые митогенетические лучи, представляющие собою ультрафиолетовые лучи короткой длины волны, продуцируемые организмами и могущие стимулировать клеточные деления в других организмах; по вопросу о митогенетическом излучении имеется уже обширная литература, и многочисленные исследователи занимаются им у нас и за границей. Метаморфоз, особенно амфибий, давно уже является объектом экспериментальных исследований; выяснено, что метаморфоз зависит от деятельности желез внутренней секреции, в особенности от щитовидной железы (Гудернач и др.), гормон которой стимулирует превращение. Процессы линьки птиц и млекопитающих изучаются с точки зрения зависимости их от гормональных воздействий (линька птиц под влиянием гормона щитовидной железы) и от воздействий химических веществ. Окраска животных, зависящая от пигментации покровных образований (кожи, перьев, волос), стоит в связи с температурными условиями, световыми и гормональными влияниями. Если у горностаевого кролика выщипать или выбрить волосы в том месте тела, где шерсть белая, и поместить затем животное в пониженную температуру, то новые волосы вырастают черного цвета (В. Шульц и др.). Гормон подмозговой железы гипофиза — влияет на окраску кожи амфибий; после удаления гипофиза наступает побледнение окраски вследствие сжатия содержащих пигмент клеток, которые со временем уменьшаются и в числе. Большие дозы гормона щитовидной железы вызывают депигментацию перьев птиц, отрастающих после линьки. Развитие признаков пола также является предметом изучения механики развития (см. феминизация, XLIII, 141, и раздельнополость, XXXV, 477/82).
Обширной главой механики развития является учение о регенерации (см.). В течение развития особи регенерация осуществляется тем легче, чем моложе стадия; у высших животных (членистоногие, высшие хордальные) способность к регенерации значительно более ограничена, чем у более низко организованных (многие кишечнополостные и черви, иглокожие, оболочники), но простой связи между высотой организации и регенеративной способностью все же нет; весьма низко организованные губки обнаруживают малую способность к регенерации; близко стоящие в системе малощетинковые кольчатые черви и пиявки весьма различны в отношении регенерации, сильно развитой у первых и весьма слабой у вторых; гребневики из кишечнополостных обладают небольшой регенерационной способностью. Установлено влияние нервной системы на процесс регенерации. Так, отрезанный передний конец у земляного червя не регенерирует на своем месте, если нервную цепочку удалить на некотором протяжении (Морган); регенерация конечностей у амфибий невозможна в отсутствии нерва. Одновременная регенерация в нескольких местах (например, более чем одного луча у морской звезды, двух ран на хвосте одного головастика) происходит быстрее, чем в одном месте.
Наконец, процессы развития особи зависят и от внешних условий, и эта сторона вопроса является предметом изучения механики развития. В силу наличия у особи наследственных факторов и вследствие того, что внутренние факторы развития связаны в систему, саморазвитие, независимость выступает здесь с особой резкостью; но детерминация направления развития возможна и здесь извне, как о том свидетельствует, например, развитие из яйца пчелы и матки и рабочей особи.
Литература: 1) Учебники и сводки по эмбриологии: К. Н. Давыдов, «Курс эмбриологии беспозвоночных» (1914); А. Заварзин, «Краткое руководство по эмбриологии человека и позвоночных животных» (1929); Г. Г. Щеголев, «Краткий курс эмбриологии человека» (1926); С. Dawidoff, «Traité d’embryologie comparée des invertebrés» (Paris, 1928); О. Hertwig, «Lehrbuch der Entwicklungsgeschichte des Menschen und der Wirbeltiere» (Jena, 1915); Corning, «Lehrbuch der Entwicklungsgeschichte des Menschen»« (1921). 2) История эмбриологии: А. Д. Некрасов, «Оплодотворение в животном царстве» (1930): Burckhardt, «Geschichte der Zoologie» (Lpz., 1921); Carus, «Geschichte der Zoologie» (Münch., 1872); Nordenskiöld, «Geschichte der Biologie» (Jena, 1926); Radl, «Geschiclite der biologischen Theorien» (Lpz., 1913). 3) Теория рекапитуляции: Бэр, «Избранные работы» (перевод с примечаниями Ю. А. Филипченко, 1924); Фриц Мюллер, «За Дарвина» (перевод с примечаниями И. И. Ежикова, 1932); А. Н. Северцов, «Этюды по теории эволюции» (1912): V. Franz, «Ontogenie und Phylogenie» (Berl., 1927); De Beer, «Embryology and Evolution» (Oxford, 1930); 4) Э. Конклин, «Наследственность и среда» (1928); М. М. Завадовский, «Динамика развития организма» (1931); Ю. А. Филипченко, «Экспериментальная зоология» (1932); П. Трендленбург, «Гормоны» (т. I, 1932); А. Г. Гурвич, «Митогенетическое излучение» (1932); Н. Przibram, «Еxperimental Zoologie» (Bd. I-V, Lpz.-Wien, 1907-1914); Т. Morgan, «Experimental Embryology» (New-York, 1927); В. Dürken, «Lehrbuch der Experimentalzoologie» (Brl., 1928); W. Schleip, «Die Determination der Primitiventwicklung» (Lpz., 1929); Е. Korschelt, «Regeneration und Transplantation» (В. I, Brl., 1927).
И. Ежиков.
| Номер тома | 53 |
| Номер (-а) страницы | 559 |