Фитопалеонтология
Фитопалеонтология, или палеоботаника (учение об ископаемых растениях), описание и изучение остатков ископаемых растений, приведение их в систему и изучение закономерностей распределения их во времени и в пространстве. Ботанику она освещает историю и ход эволюции растительного мира, геологу служит: а) для определения возраста геологических отложений (стратиграфическое значение ископаемых растений); б) для освещения условий, в которых протекала жизнь в отдельных участках земной поверхности (экология ископаемых растений), и в) для выяснения изменения конфигурации земной поверхности в минувшие геологические времена (ареалы распространения ископаемых растений).
Ископаемые растения крайне редко доходят до нас в полном виде с сохранением всех своих частей, которые давали бы нам материал для суждения об их внешних признаках и внутреннем строении. Чаще всего они доходят до нас в виде фрагментов: отдельных кусков стеблей, черешков, корней, разобщенных листьев или их частей, отдельных плодов или органов размножения в разных степенях сохранности. Они могут дойти до нас в разных состояниях: или 1) в минерализированном состоянии с сохранением всех деталей внешнего и внутреннего строения, например, в окремнелом или доломитизированном состоянии; или 2) в виде отпечатков, позитивных или негативных, наружных частей растений — листьев, стеблей, корней, плодов, семян и т. д., перешедших в ископаемое состояние с неизмененными или измененными поверхностями; или 3) в виде слепков с пустот, которые были выполнены телом растения и оказались заполненными осадочной породой после смерти и разложения растения; или, наконец, 4) в виде пустот в различных горных породах, которые после заполнения их в лабораториях расплавленным воском или другими веществами могут дать муляжи тех растений, которые исчезли из этих пустот с течением времени. Наибольшую научную ценность представляют ископаемые растения первого рода. Приготовляя шлифы из таких остатков, можно под микроскопом изучать мельчайшие детали анатомического их строения. Однако, и отпечатки, особенно листьев, можно использовать в целях микроскопического изучения, пользуясь методом получения «коллоидальных оттисков» с листовых и других поверхностей ископаемых растений, методом, разработанным палеоботаником Натгорстом. «Коллоидальные оттиски» при боковом освещении под микроскопом дают поразительно ясные изображения границ клеток кожицы, устьиц, волосков и т. д. Наконец, о существовании некоторых ископаемых растений мы можем судить по тем горным породам, которые созданы или 1) скоплением их скелетов (диатомовый трепел), или 2) их выделениями (янтарь — смола третичных хвойных, известняки альконского возраста — продукт жизнедеятельности сине-зеленых водорослей, известняки триаса Альп — известковых водорослей, кораллиней), или 3) продуктами их разрушения (каменный уголь, торф, горючие сланцы, сапропель).
Изучение ископаемых растений представляет целый ряд затруднений. Во-первых, при наличности большого числа случаев конвергенции как во внешних, так и во внутренних признаках, суждение по аналогии с ныне живущими формами может быть иногда неосторожным. С другой стороны — ввиду того, что по правилам бинарной номенклатуры каждое ископаемое растение или, точнее, фрагмент с определенными признаками обыкновенно обозначается соответствующим родовым и видовым именем, может случиться, что ископаемые остатки, принадлежащие одному и тому же виду растения, могут носить разные родовые названия. Например, у семенного папоротника Lyginopteris Oldhamia были описаны листья в качестве Sphenopteris Ноеninghausii, черешки — в качестве Rahiopteris aspera, корни — Kaloxylon Hookeri, семена — Lagenostoma Lomxi, микроспорофиллы — Crossotheca Hoeninghausi и стебель с сохранившимся анатомическим строением — Lyginodendron Oldhamium. Сохранение всех этих отдельных названий, однако, необходимо для использования фитопалеонтологического материала в целях стратиграфических. Одними из первых, признавших в ископаемых растениях остатки действительно когда-то существовавших растений, были Леонардо да Винчи и Палисси. Первые работы по фитопалеонтологии носили характер описания ископаемых растений, которое сопровождалось рисунками ископаемых образцов. Таковыми надо считать «Допотопный гербарий» — Herbarium diluvianum — Шейхцера (1709), «Ископаемая Силезия» — Волкманна (1720), «Описание замечательных отпечатков трав» — Шлотгейма (1804), «Опыт изображения флоры прошлых веков» — Штернберга (1820) и др. Первой блестящей попыткой научной обработки фитопалеонтологического материала являются работы А. Броньяра: «Классификация и распространение ископаемых растений» (1822), «Введение в историю ископаемых растений» (1828) «История ископаемых растений» (1828—1838) и «Таблица родов ископаемых растений». Вскоре после появления в свет первых работ Броньяра появилась знаменитая «Ископаемая флора Великобритании», совместная работа ботаника Линдлея и геолога Хёттона (1831—1837). Огромные услуги фитопалеонтологии оказали описанием целого ряда ископаемых флор из иностранцев: Геер, Лескерё, Циньо, Массалонго, Сапорта, Эттингаузен, Гёпперт, Натгорст, Сьюорд, и из русских: Шмальгаузен, Эйхвальд, Траутшольд, Карпинский, Залесский, Криштофович, Палибин и др. Параллельно с описанием ископаемых флор начинается по стопам А. Броньяра изучение и анатомического строения ископаемых растений. Особенно велики в этом отношении заслуги английской и французской школ палеоботаников. Фитопалеонтология, ставящая своей задачей освещение истории и хода эволюции растительного мира, в первую очередь должна стремиться найти исчезнувшие звенья из той мозаичной картины, каковой рисуется нам система растительного мира. Открытие Кидстоном и Лангом в лице класса Psilophytales — псилофитов — древнейших сухопутных растений из девона, предков кормофитных (побегоносных) растений; открытие Bertrand, Renault и др. группы первичных папоротников, объясняющих стадии эволюции побега; открытие Скоттом и его школой группы семенных папоротников (Pteridospermeae), которые объединяют низшие кормофитные формы с высшими (папоротниковые с семенными); открытие Виландом и его последователями органов размножения у мезозойских саговниковых (Сусаdophyta), конвергентно осуществивших цветок высших цветковых растений, — эти выдающиеся факты, заполняя угасшие звенья растительного мира, объясняют нам в то же время пути эволюции и предостерегают от чересчур поспешных и неверных а prior’ных и чересчур примитивных представлений о ходе эволюции. Открытия эти оказали огромное влияние на современные представления о системе растительного мира, до сих пор строившиеся исключительно на основании изучения лишь ныне живущих растений.
Первый вывод, к которому приводит фитопалеонтология, это факт постепенного усложнения и прогрессивной дифференциации как во внутреннем строении, так и во внешнем габитусе друг за другом следовавших групп. Родиной растений была водная среда. Постепенно эта первоначальная арена жизни расширялась за счет суши; в этом видят причину различия между низшими — слоевцовыми клеточными растениями (Thallophyta) и побегоносными высшими, обладающими дифференцированными на ткани клетками (Cormophyta).
Второй принцип «адаптивной радиации», иллюстрируемый фитопалеонтологией, состоит в том, что каждая новая группа растений, выступавшая на арену геологической жизни [например, Lepidopliyta или Lycopsida, Arthrophyta или Sphenopsida и Pteridospermeae в палеозое, Сусаdophyta в мезозое, покрытосеменные (Angiospermae) в кенозое], захватывала господствующее положение благодаря тому, что ее представители с помощью прогрессивных мутаций становились приспособленными к огромному разнообразию условий среды и стремились захватить все доступные местообитания на суше, не останавливаясь даже перед вторичным приспособлением к жизни в водной среде (водяные папоротники и различные водяные покрытосеменные). Прилагаемая схема Берри иллюстрирует последовательное господство различных типов растений и постепенное возрастание сложности состава растительного мира в целом (рис. 1.).
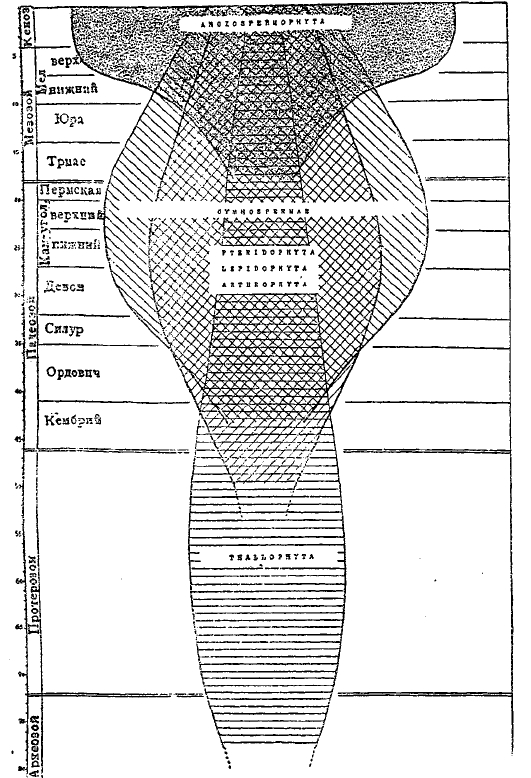
Рис. 1. Относительная продолжительность геологического времени, последовательное преобладание растительных типов и прогрессивное осложнение в составе флоры.
Третий принцип фитопалеонтологии заключается в том, что по мере того, как организм или органы становятся все более и более сложными и все более и более специализированными, — прогрессивно уменьшается их пластичность. Организмы с более простым строением предшествуют более сложным и дают начало новым типам. Более сложные организмы утрачивают свою способность приспособляться к изменениям среды и легко гибнут. Этому положению противоречит как будто факт существования в современной флоре хвощей и плаунов, но на деле, не сложно построенные древовидные лепидодендроны дали начало современным плауновым, и не высокие ростом каламиты выродились в современные хвощи, а в каждом из этих случаев более простые формы спасли данную родословную ветвь от окончательного уничтожения. Синангии папоротников (см. XXXI, 132) путем видоизменения и стерилизации периферических спорангнев дали начало семени, но семя никогда не превращалось и не превратится в синангий. Эволюция каждого органа или организма необратима. У более ранних форм следующих друг за другом групп растений, в полном соответствии с а prior’ными заключениями эволюционной философии, скомбинированы признаки разных категорий, которые в дальнейшем становятся характерными для разных расходящихся линий эволюции. Так, например, наиболее древние папоротники — Primofilices — обнаруживают комбинацию признаков, которые в дальнейшем становятся характерными признаками различных семейств папоротников; равным образом и семенные папоротники комбинируют признаки папоротников и Cycadophyta.
Наиболее консервативными органами являются органы вроде корней, у которых и функции и среда остались единообразными в течение веков. Поэтому неудивительно, что все корни, как у ископаемых, так и у ныне живущих растений весьма схожи в чертах своего строения. Листья, в общем, тоже склонны к сохранению, как внешней формы, так и внутреннего строения, что подтверждается сходством в строении листьев в группе Cycadophyta, сохранившемся неизменным от пермского периода до наших дней, тогда как за это время сосудистое строение стебля и особенно их органы плодоношения изменялись в широких пределах.
Наконец, еще один весьма важный вывод мы получаем из изучения распределения растений в течение геологического времени. Он касается геологического климата, наиболее надежными свидетелями которого могут быть ископаемые растения. Начиная с нижнего девона, времени, к которому относится открытие первых сухопутных растений, до настоящего времени было только два периода, в которые возможно различить сколько-нибудь приблизительную климатическую зональность. Так, например, девонские растения, описанные из отложений Ellesmere Land на севере, Австралии на юге и центральной Европы не обнаруживают никаких различий. Такое единообразие и космополитизм в распространении продолжается через весь нижний и верхний карбон. Правда, в пермский период существовали хорошо разграниченные флористические провинции в результате широко распространенных в то время явлений оледенения. Но вслед затем, начиная с раннего триаса, через юру и мел, мы опять находим одни и те же растения как в нескольких градусах от полюса, так и у экватора. Даже в первой половине третичного периода тропическая растительность простиралась до середины теперешнего умеренного пояса, а леса умеренной зоны известны из мест, расположенных в 5° от полюса. Наступившее к концу третичного периода оледенение в плейстоцене нарушило весь этот космополитический строй, и с того времени и до наших дней человечество живет в плейстоценовом климате, с ясно различимыми климатическими зонами и изменениями по сезонам.
Относительно первых стадий развития растений на земле приходится ограничиваться теоретическими предположениями и заключениями. Открытия в области бактериологии показали, что первыми живым организмами на земле могли быть организмы, которые все свое питание строили исключительно на использовании неорганических материалов, и таким образом представляли до возникновения хлорофилла первую стадию эволюции жизни на земле. Современные чисто прототрофные бактерии иллюстрируют нам способы, какими в этой дохлорофилловой стадии простые организмы добывали себе необходимую для осуществления жизненных процессов энергию путем окисления углерода, азота, серы или железа, которые они получают из сероводорода, аммиака и т. д. (см. хемосинтез). Таким образом, исторически хемосинтез должен был предшествовать фотосинтезу. Современные научные данные показывают, что некоторые формы фотосинтеза, при помощи которых определенные бактерии используют пурпурный и другие пигменты для утилизации инфракрасных лучей света, весьма вероятно предшествовали и могли быть ступенью к образованию хлорофилла.
Благодаря современным открытиям в области ископаемых бактерий признано, что денудационные процессы поверхностей наиболее древних горных пород осуществлялись при содействии бактерий, которые способствовали выветриванию пород такими же способами, как это они делают в настоящее время. Следующая стадия в истории эволюции бактерий, представленная в настоящее время метатрофными формами, состояла в том, что эти бактерии вместо того, чтобы добывать необходимый для своей жизнедеятельности углерод из неорганических источников, стали его добывать из сложных азотистых соединений других бактерий или водорослей, как только последние появились. Третья стадия в истории бактерий, представленная в настоящее время паратрофными бактериями, наступила тогда, когда они перешли к настоящему паразитическому образу жизни. Эволюция метатрофных бактерий, очевидно, стала осуществляться вблизи того периода истории земли, в котором стал возможен процесс созидания сложных органических молекул из неорганических материалов; с этого времени деятельность бактерий стремится задержать этот процесс, и в этом обстоятельстве, между прочим, можно искать причину того, что возникновению жизни в более поздние геологические времена был положен предел.
Докембрийские известняки, равно как известняки более позднего времени, в которых мы не находим никаких определенных следов организмов, теперь рассматриваются геологами, как указания на существование бактерий в качестве активных агентов отложения, подобно тому, как это имеет место в настоящее время при осаждении современного ила, содержащего известь. Равным образом железные бактерии, вероятно, участвовали в процессах отложения железных руд в первичных породах (ср. железо и железоделательное производство, XX, прил. к ст. 152, 12/13).
Присутствие протерозойных бактерий было доказано работами Walcott’а. Палеозойские бактерии впервые были открыты в 1879 г. Ван-Тигемом, который исходил из доказанных следов масляно-кислого брожения клетчатковых оболочек у окремневших впоследствии растений из Saint-Etienne. Вслед за тем значительное число ископаемых палеозойских бактерий было описано Рено и другими палеоботаниками путем микроскопического изучения шлифов ископаемых растений и копролитов (окаменевших выделений животных). Доказательства деятельности бактерий очевидны на материале из окременелых растений всех геологических возрастов. Историю эволюции растительного мира правильно было бы разделить на две стадии. Дохлорофилловая стадия, приведшая, в конце концов, к образованию наиболее простых форм водорослей, как будто потребовала времени не меньше, чем вторая — хлорофилловая, обнимающая всю дальнейшую эволюцию растительного мира. Но если принять во внимание, что протоплазма этих одноклеточных форм находилась в непосредственном соприкосновении с окружающей средой, и что влияние окружающей среды было неизмеримо сильнее, чем после того, как протоплазма оказалась заключенной в оболочку из целлюлозы и стала относительно более инертной благодаря образованию специальных систем тканей, тогда можно допустить, что для этой первой стадии эволюции не потребуется такого количества времени, как это представлялось необходимым до сих пор. В течение первой дохлорофилловой стадии история растительного мира постепенно переходила во вторую, или хлорофилловую фазу, которую можно назвать фазой эволюции водорослей. Первоначальные бактериеподобные формы жизни получали необходимую для жизни энергию из геосферы и гидросферы. Образование хлорофилла сделало их способными к использованию атмосферы, как источника энергии. Это развитие хлорофилла было, может быть, величайшим шагом вперед в эволюции жизни, вторым по своей важности после возникновения жизни вообще. Эта вторая фаза эволюции растительного мира началась весьма давно; это обнаруживается следами водорослей в отложениях докембрийских и целым рядом косвенных доказательств, свидетельствующих об активности растительного мира, вроде, например, огромных скоплений углерода (графита) в протерозойных отложениях, вроде присутствия скоплений кремнезема в виде опалов, обусловленных деятельностью водорослей, и т. п.
Ближайшими к бактериям являются сине-зеленые водоросли (см.). Ядро их клетки, по-видимому, не имеет оболочки, их хлорофилл не сосредоточен окончательно в хлоропластах, и размножаются они исключительно путем дробления. Водоросли стали дифференцироваться как в своей вегетативной, так и в половой сфере и приспосабливаться к различным местообитаниям в море и изменяться, но, в общем, очень слабо в течение геологического времени. Среди наиболее примитивных зеленых водорослей и бурых водорослей мы находим немало представителей той ближайшей ступени в эволюции растений, которая закончилась вторжением их на сушу. Последнее привело, в конце концов, к созданию разнообразной древесной растительности с плодами и цветками, как это мы видим в настоящее время. Этой последней стадии в истории растительного мира, которая началась в додевонские, а весьма вероятно и докембрийские времена и которую можно назвать стадией кормофитных (стеблевых) растений, предшествовала некоторая промежуточная стадия, которую можно назвать стадией прокормофитных растений, или псилофитов. Представители водной среды успели уже выработать внутреннюю анатомическую дифференцировку, приспособленную к жизни в воздушной среде, но не успели еще выработать внешнее расчленение на стебель, корень и лист. Эта промежуточная стадия, последние представители которой вымерли к середине девона, сменилась окончательно последней, кормофитной стадией, с начальных моментов которой берут свое начало не только крупнолистные папоротники, семенные папоротники, цикадовые, хвойные и цветковые растения, но и микрофильные лепидофиты и артрофиты; надо заметить, что мохообразные (Bryophyta) представляют независимую и никогда не бывшую всецело сухопутной линию эволюции, которая берет свое начало от других типов водорослей и, таким образом, никогда не находилась в непосредственном родстве с другими высшими растениями.
Несомненно, наиболее важными моментами в истории растительного мира были первые овладевания суши. Несомненно, мы должны смотреть на водоросли, как на группы, давшие начало сухопутным растениям; это следует из земноводного способа оплодотворения так называемых архегониат.
Весьма вероятно, что некоторые из зеленых и бурых водорослей первыми покинули водную среду. Потребность в воде привела в результате к образованию органов для восприятия воды (корни) и к образованию кутикуляризированного эпидермиса для ограничения траты воды, а затем неизбежно к эволюции механической ткани и к образованию проводящих тканей, в результате чего растения оказались способными приподнять свои органы ассимиляции над поверхностью почвы.
Способность к образованию вторичной древесины у различных ветвей наиболее древних из сухопутных растений явилась для них большим преимуществом и имела важные последствия. Нарастание в диаметре повело к достижению большей высоты и к более сильному ветвлению; последнее привело к увеличению ассимилирующих тканей и к более совершенным проводящим тканям. У некоторых линий эволюционного развития растительного мира, в роде, например, кордаитов, развитие стебля шло по пути, по которому идет развитие у современных деревьев. Другие линии пытались строить стебель с помощью центростремительной в своем развитии вторичной древесины, которую мы находим у древних тайнобрачных. Третья линия эволюции - лепидодендроны, смогла осуществить высокий и стройный рост, ограничившись только тонкой зоной вторичной древесины, возложив все механические задачи на образование толстой коры, но разрешение задачи оказалось неудачным, и они исчезли вместе с палеозоем.
Весьма важным приспособлением с точки зрения эволюции, возникшим очень рано, оказалось образование гетероспории в таких родословных линиях, которые первоначально имели споры однородного типа. Благодаря развитию двух родов спор, макроспоры оказались лучше приспособленными в борьбе за существование, а микроспоры должны были образовываться в значительно большем числе, чтобы таким путем увеличить шансы оплодотворения. Следующий шаг — удерживание макроспоры в теле материнского растения на все более и более продолжительный период, в течение которого материнское растение снабжает ее нужною пищей и водой и развивает аппарат, облегчающий доступ сперматозоиду к яйцеклетке. Этот шаг привел к образованию настоящих семян, к созданию которых почти приблизились лепидофиты и осуществили семенные папоротники. Растения, обладавшие семенами подобно живородным млекопитающим, оказались господствующим классом в настоящее время, что очень явственно подчеркивает преимущества семени, как органа размножения.
Ископаемые растения могут быть систематизированы в следующие группы. Слоевцовые, обнимающие собой водоросли и грибы; прокормофитные, или псилофиты, по своим признакам занимающие промежуточное место между слоевцовыми и следующей группой — кормофитных растений. Последняя группа до последнего времени разделялась на папоротникообразных (Pteridophyta), голосеменных (Gymnospermae) и покрытосеменных (Angiospermae).
Из водорослей дошли до нас по преимуществу такие формы, которые способны были еще заживо создать свой скелет и отложить вокруг себя минеральные вещества. Такими являются сине-зеленые водоросли Collenia, Newlandia, Greysonia и др. из известковых отложений альконского возраста; диатомовые водоросли, скелеты которых, образующие скопления трепела, горной муки и т. д., встречаются, начиная с нижней юры; из зеленых водорослей оставили после себя несомненные следы представители сифонниковых (Sicydium, Ascoma). Они встречаются от кембрия до силура, но особенно часто (Diploporella, Triploporella и др.) образуют известняки триаса Альп и отложения миоцена парижского бассейна; багряные водоросли (Solenopora) начинают попадаться в ордовичских (нижне-силурийских) слоях и особенно (Lithothamnion и Lithophyllum) в известняках мезозоя и кенозоя. Также обычны ископаемые лучицы, споропочки и осколки стебельков которых встречаются, начиная с юры, а, по мнению Карпинского и в девоне России. Бурые водоросли встречаются также, начиная от силура (Nemathophycus) и дальше до отложений ледниковых в виде отпечатков, напоминающих по внешности современные Fucus и др. Из низших растений в ископаемом состоянии известны грибы, или в виде гиф грибного мицелия, пронизывавших древесины ископаемых деревьев, или в виде следов органов плодоношения, отпечатавшихся на поверхностях ископаемых листьев.
Самой интересной группой из ископаемых растений в настоящее время считается группа псилофитов (Psilophytales), установленная только в 1917—1921 г. (роды: Rhynia, Hornea, Asteroxylon, Psilophyton и др.). Эти, несомненно, древнейшие (с нижнего девона) из известных нам сухопутных растений представляют собой формы самые примитивные среди высших сосудистых сухопутных растений. Характерными признаками их были наличность расчленения клеток на ткани: так, у них мы находим эпидермис, устьица, механическую ткань, ассимиляционную ткань и сосудисто-волокнистый пучок — стелю, занимающую центральное положение в оси тела растения и состоящую исключительно из элементов первичной ксилемы. При таком сравнительно высоком уровне внутреннего строения растение отличалось необычайной внешней простотой. Тело растения, например, у Rhynia, представляло лишь дихотомически разветвляющуюся ось без всяких намеков на расчленение на основные органы высших кормофитных растений: корень, стебель и лист. При наличности такой простоты внешнего строения, тем не менее существовало уже различие между участками оси, обслуживавшими потребности индивидуальной жизни (трофические), и участками, обслуживавшими потребности видовой жизни (спорогенные). Замечательно, что спорангии могли, например у Hornea, дихотомически ветвиться. Открытие этих примитивных сухопутных растений, связующих в своих признаках низшие слоевцовые с высшими побегоносными (кормофитными) растениями, бросает совершенно новый свет на эволюцию сухопутной флоры. В связи с этим можно рассматривать мохообразные (Bryophyta) не как предков высших растений, а как обособленную ветвь растительного мира, возникшую может быть путем регрессивной эволюции из какой-либо высшей группы, за что говорит появление в ископаемом состоянии несомненных мхов лишь начиная с мела.
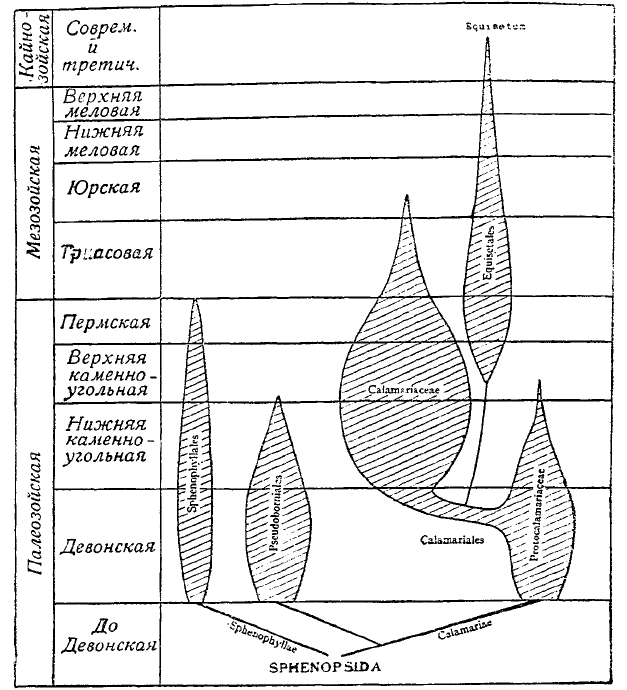
Рис. 2. Геологическая история и филогения Sphenopsida.
Фитопалеонтология, давшая колоссальный материал по истории развития сухопутной флоры, изменяет наши представления о системе растительного мира, которые сложились на основании изучения, главным образом, ныне живущих растений. Деление высших кормофитных (побегоносных) растений на папоротникообразные, голосеменные и покрытосеменные, в основу которого положены были, главным образом, данные эмбриологии, носит в себе элементы искусственной системы, поскольку принят был во внимание лишь один принцип (смена поколений). Если же, кроме данных эмбриологии, принять во внимание данные морфологии спорофита, данные строения стели (оборудования стебля сосудистой системой, характер строения сперматозоидов и т. д.), то придется признать, как это думает Скотт и целый ряд других палеоботаников, что сухопутная флора прошлого и настоящего представляет три, а может быть даже четыре самостоятельные эволюционные ветви, корнями своими уходящие в морские водоросли. Ветви эти крупнейший из современных палеоботаников, Скотт, называет так: 1) Sphenopsida или Articulatae (Arthrophyta) — или растения с членистыми побегами; 2) Lyeopsida или Lepidophyta — растения плауноподобные, и 3) Pteropsida — растения папоротникового колена растительного мира, которое Скотт подразделяет по следующей схеме:

Последние три группы Spermophyta (семенные растения) Скотт считает даже возможным рассматривать, как самостоятельную линию эволюции, независимую от собственно папоротников, на том основании, что в среднем девоне доказано существование стволов высокоорганизованных семенных папоротников (Pteridospermeae) или голосеменных (например, Palaeopitys Milleri), тогда как у нас нет никаких несомненных доказательств существования в начале девона настоящих папоротников.
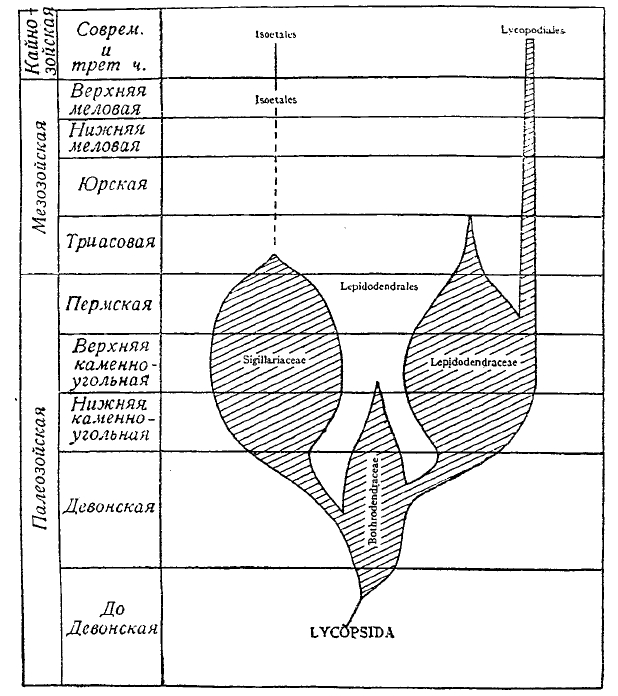
Рис. 3. Геологическая история и филогения Lyсopsida.
Sphenopsida (Arthrophyta) обнимают собой формы, впервые появляющиеся в ископаемом состоянии в девоне в лице первичных каламитов (Protocalamiteae), псевдоборний (Pseudohomiales) и клинолистов (Sphenophyliales) — отличавшихся членистыми стеблями, сложно-дихотомически рассеченными листьями, расположенными мутовками, и спорофиллами, собранными в верхушечные колоски. На смену первым двум в карбоне появляются каламиты (Calamites) с простыми листьями и стеблями, в виде высоких деревьев, обладавших способностью ко вторичному утолщению стебля. От этих растений дошли до нас отпечатки листьев, описанные под родовыми названиями Annularia, Asterophyllites; органы плодоношения в виде спороносных колосков, описанные под названием Calamostachys, Palaeostachya, Macrostachya; слепки с внутренних полостей стебля с характерной наружной скульптурой, описанные под названием Calamites, Calamitea; участки стеблей с окремнелыми частями стебля Calamodendroa, Arthropitys, Arthrodendron; корни с сохранившимся внутренним анатомическим строением — Astromyelon. Каламиты, бывшие характерными деревьями в течение всего карбона и пермского периода, начиная с триаса, понемногу сходят со сцены и исчезают к началу юры. Почти одновременно, но совершенно независимо обнаруживается веточка этого колена растительного мира — хвощовые, которые, достигши максимума своего развития к концу пермского периода и к началу триаса, начиная с юры стали уменьшаться и доходят до наших дней в виде одного семейства, содержащего только один род с 25 видами. Характерно то, что данные фитопалеонтологии рисуют нам эволюцию этой ветви растительного мира не с самого начала, а как бы с апогея их морфологического развития. Вся дальнейшая геологическая история этой ветви это — история последовательного падения (см. рис. 2).
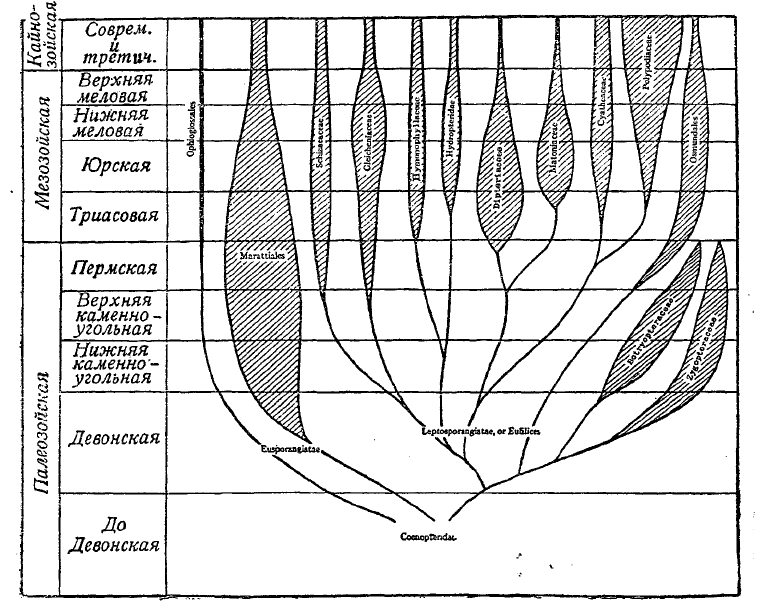
Рис. 4. Геологическая история и взаимоотношения папоротников.
Lycopsida (Lepidophyta) впервые появляются в верхнем девоне. Один из представителей этой группы Protolepidodendron primaevum, найденный в окрестностях Нью-Йорка в Северной Америке, соединяет в себе целый ряд признаков, которыми характеризовались остальные представители этой ветви ископаемых растений: семейство Bothrodendraceae (верхний девон и нижний карбон), семейство Sigillariaceae и Lepidodendraceae (весь карбон и пермь) (см. рис. 3). Эти два вымерших семейства оставили в живых лишь три небольших группы своих потомков, в лице современных травянистых семейств Isoetaceae, Lycopodiaceae и Selaginellaceae (см. плаунные). О строении ископаемых растений этой ветви могут дать наиболее полное представление так называемые чешуедревы — или лепидодендры (см. плаунные). Сигиллярии (см.) и лепидодендроны, несомненно, были обитателями болотистых почв, на что указывает сильное развитие как в стеблях, так и листовых подушечках, воздухоносных полостей и воздухопроводящих тканей. Участки стеблей, сохранившие внутреннее анатомическое строение, свидетельствуют о способности стеблей лепидодендронов к образованию вторичной древесины и утолщению стебля. Обилие воздухоносных полостей в стеблях сигиллярий и лепидодендронов, ксерофитная структура их листьев и т. д. бросают свет на условия местообитания и их экологию. Несомненно, что эти растения жили в условиях физиологической сухости субстрата вроде тех, в которых в настоящее время живут в тропических странах мангровые.
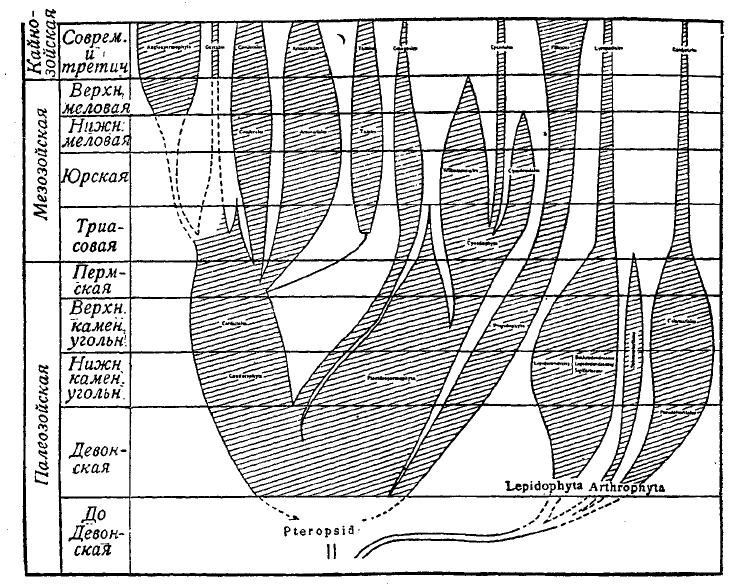
Рис. 5. Геологическая история и филогения сосудистых растений.
Pteropsida. Сюда входят, по Скотту, Fiticales, т. е. собственно папоротники, как первичные (Primofilices), так и все остальные (см. рис. 4), и Spermophyfca, т. е. семенные растения, начиная от так называемых семенных папоротников (Pteridospermeae), цикадовых-беннетитов, кордаитов из числа ископаемых голосеменных и тех голосеменных и покрытосеменных, которые составляют главную массу современной флоры (см. рис. 5). Древнейшей группой среди ископаемых папоротников является группа так называемых первичных папоротников (Primofilices), представленных двумя семействами Zygopteridaceae и Botryopteridaceae. Формы эта обладали стеблем с одной центральной стеблей. На этих стеблях находились вайи очень простого строения. Скорее всего, это были оси, заканчивавшиеся на вершине, как у псилофитов, спорангиями, сидевшими кучками по 4—8 спорангиев, которые могли быть даже объединены в синангий. На этих вайях-осях по сторонам находились слабовыраженные крыловидные пластинки, зачаточные ассимиляционные листовые поверхности. Primofilices существовали в течение палеозоя и с концом пермского периода вымирают. Их современниками было семейство Marattiaceае, некоторые роды которого (в числе 6) дожили до наших дней и принадлежат флоре палеотропических и неотропических стран. К представителям палеозоя из этой группы папоротников должны быть отнесены некоторые отпечатки листьев с сохранившимися следами спорангиев, собранных в синангиальные сорусы, а целая группа так называемых Psaroniaceae, дошедших до нас в виде удивительно хорошо сохранившихся стеблей с закономерно распределенными стеблями, окруженных целой мантией придаточных корней так же, как у некоторых из современных древовидных папоротников. Характерно, что Primofilices в некоторых чертах своего строения приближаются к мараттиевым, а другие к Osmundales, в группе папоротников, появляющихся вместе с семействами Schizaeaceae в Gleicheniaceae начиная с пермского периода, достигших максимума своего развития в середине мезозоя и затем пошедших на убыль, но встречающихся в лице немногих представителей среди современной нам флоры. Эти три семейства характеризуют уже флору мезозоя. Но наиболее характерными папоротниками для мезозоя надо считать два семейства Dipteriaceae и Mattoаіасеае, ставшие вымирать, начиная с верхнего мела, и уцелевшие в настоящее время лишь в одной области по обоим сторонам Малаккского пролива. Начиная с юры можно проследить появление представителей семейства Polypodiaceae, которое в настоящее время находится в апогее своего развития, и изучение которого кладет свой преимущественный отпечаток на наши познания о папоротниках. Ввиду обилия отпечатков листьев типа папоротников, которые раньше относили к мараттиевым, в отложениях каменноугольной и пермской систем, флору карбона и перми раньше характеризовали, как флору папоротникообразных. Однако, еще в начале 80-х годов XIX ст. австрийский палеоботаник Weiss высказал сомнение в принадлежности их к папоротникам. Через двадцать, приблизительно, лет Скотт вместе со своим учеником Оливером выступили с докладом о так называемых семенных папоротниках (Pteridospermeae), т. е. о растениях, у которых на вайях непосредственно сидели семена, и которые, таким образом, соединяли в себе одновременно признаки папоротников и семенных растений. Открытие Скотта естественно вытекало из исследований Williamson'а, касавшихся анатомических особенностей целого ряда ископаемых стеблей, черешков, листьев и корней различных палеозойских растений, описанных под разными названиями Lyginodendron Oldhamium, Rhachiopteris aspera, Kaioxylon Hookeri. Все они оказались частями одного и того же растения. Семена были построены по типу близкому к семенам цикадовых, имели в нуцеллярном носике пыльцевую камеру, в которой, вероятно, плавали подвижные сперматозоиды, как это можно было заключить по некоторым деталям строения найденных в пыльцевой камере микроспор. Таким образом, удалось реконструировать целое растение, отнесенное в роду Lyginopteris. По мнению Скотта, это было высокое растение типа лазающих лиан, жившее в палеозойских лесах карбона и перми, снабженное крупными вайями в виде двух симметричных половинок, сидевших на раздвоенном в верхней своей части черешке. Вскоре ряду исследователей удалось открыть целую серию родов, близких к Lyglnopteris, и установить параллельное Lyginopteridaceae семейство Medullоsасеае. Открытие группы семенных папоротников имело громадное значение: 1) оно показало, что все прежде бывшие прямолинейные представления об эволюции семян не соответствуют действительности, 2) что семенные растения так же древни, как споровые кормофитные, так как встречаются, начиная с верхнего девона и 3) нахождение семяпочек на концах черешков ваий говорит в пользу осевого происхождения семяпочки, так как вайи папоротников рассматривают, как разветвленную систему осей, объединенных в целях ассимиляции в плоские листовидные органы, а каждый спорангий — как естественное завершение концов таких осей, что в свою очередь совпадает с тем, что дают нам псилофиты.
Семенные папоротники, по существу своему могущие быть названными древнейшей группой голосеменных, являются характерными растениями палеозоя. Другой ветвью палеозойских голосеменных являются кордаиты (см.). Pteridospermeae и Cordaitales исчезают с конца триаса, а на смену им в мезозое появляются связанные с ними две новые группы форм, одна, берущая начало от Pteridospermeae, а другая — от Cordaitales. К первой группе, которую можно назвать мезозойскими Cycadophyta, или Веnnеtitales, относятся два порядка: Cycadeoidales и Williamsoniales, оба вымершие: первый к концу верхнего, а второй к концу нижнего мела, и третий порядок Cycadales, немногие представители которого дожили до наших дней, но характеризуются ясно выраженным эндемизмом в пределах тропиков Нового и Старого Света. Ко второй группе голосеменных относятся хвойные (Coniferae).
Наибольший интерес представляют Bennetitales. Ископаемые стволы их известны под названиями: Cycadeoidea и Bennetites. Эти растения имели толстые клубневидные стволы, от нескольких см, до 8—4 м высотой и от нескольких см до 1,5—2 метров в диаметре. Поверхность стволов покрыта была броней из остающихся оснований листовых черешков, как это наблюдается у многих из современных цикадовых. Эти основания листовых черешков были погружены в густую массу чешуй, покрывавших поверхность стебля. У некоторых стеблей оказались сохранившимися конического вида верхушечные почки, у которых вместо листьев можно найти линейные чешуя. В листовых черешках сохранилось расположение сосудисто-волокнистых пучков, на поперечном разрезе расположенных в виде подковообразной фигуры с согнутыми внутрь краями; в основной ткани разбросаны смоляные ходы, а на поверхности сильно развита перидерма, как у современных цикадовых. Обилие чешуй, покрывавших стебель, плотное строение хорошо защищенных цветков, снабжение чешуй эпидермисом и листьев толстой кутикулой — все это указывает на ксерофильные признаки, выраженные в сильной степени. Стволы беннетитов, по-видимому, имели вторичную древесину значительной толщины с ясными признаками концентрического строения ксилемы, что могло быть результатом некоторой периодичности в остановках роста. Древесина состояла из лестничных трахеид со спиральными элементами в первичной своей части. Флоэма представляла чередование толстостенных элементов и тонких клеток. В коре из паренхиматических клеток были разбросаны широкие выделительные каналы. Обширная сердцевина состояла из паренхиматических клеток и выделительных каналов. Очень редко можно встретить листья, сохранившие связь со стеблями и описанные под названиями: Zamites, Ptilophyllum и Dictyozamites. Характерным для стволов беннетитов является нахождение особых шишек или цветков, которые погружены более или менее в броню из оснований черешков и сидят на верхушках боковых пазушных побегов, одетых у основания спирально расположенными редуцированными листьями, или прицветниками; эти линейные многочисленные прицветники покрыты густым войлоком волосков и образуют серьезную защитную оболочку. Вершина плодущего побега оканчивается закругленной полушаровидной вершиной — цветоложем. Цветки по большей части двуполые. Замечательным признаком является огромное число цветков на отдельном стебле; так, у Cycadeoidea Dartoni, длиной в 54 см и шириной в 86 см, оказались сосредоточенными на одной стороне стебля от 500 до 600 цветков. Можно думать, что растение производило громадное количество цветков после значительного времени исключительно вегетативной жизни и, подобно, например, современной нам пальме Corypha, достигши высшей ступени в своем индивидуальном развитии, умирало. В развитии цветков на старых стволах Cycadeoidea мы имеем поразительный пример каулифлории (образование групп цветков на стебле и старых ветвях). По строению цветков беннетиты резко отличались от современных цикадовых. Двуполые цветки по форме напоминают куриное яйцо или грушу средних размеров и представляют заметный контраст с мужскими или женскими шишками современных цикадовых. Мутовка перисторазветвленных микроспорофиллов прикреплялась к нижней части цветоложа в виде трубки, образованной сросшимися основаниями спорофиллов. Каждая долька перисторазветвленного микроспорофилла несет несколько синангиев на коротких ножках, подразделенных на гнезда, содержавшие микроспоры с гладкими оболочками, которые напоминали микроспоры современных цикадовых. После развертывания и высыпания спор, весь мужской диск опадал, оставляя узкий ободок внизу полушаровидного или конического цветоложа. Верхняя часть цветоложа была покрыта семяпочками, сидевшими на стебельках, вперемежку с так называемыми межсеменными чешуями, и каждый стебелек был окружен 5—6 чешуями. Семена были, по-видимому, без эндосперма и с зародышами типа двудольных и имели 8—4 мм в длину.
Стволы беннетитов были найдены в разных частях Европы, но особенно много, свыше 1000 стволов, найдено было в Северной Америке в верхнеюрских и нижнемеловых отложениях.
Весьма близким родом к Cycadeoidea был род Williamsonia, цветки которого дают возможность проследить эволюцию цветка путем постепенной редукции микроспорофиллов и превращения стерильных частей микроспорофиллов в весьма вероятные листочки околоцветника, предваряя цветов будущих покрытосеменных.
Представители родов Wielandiella и Williamsoniella обладали тонкими, ложнодихотомически ветвившимися стеблями, покрытыми листьями или спирально расположенными (Williamsoniella), или сближенными в мутовки выше развилков (Wielandiella). Из середины развилка выходило по одному цветку, который у Williamsoniella состоял из центральной оси — грушевидного цветоложа с макро- и микроспорофиллами. Макроспорангии (семяпочки) не имели стебелечка и были сидячие, микроспорофиллы окружали цветоложе и функционировали одновременно и как покровы цветка; они сидели мутовкой, каждый нес по 5-8 синангиев, содержавших микроспоры. Синангии по три были расположены по бокам уплощенной части микроспорофиллов, напоминая как бы двустороннее расположение пыльцевых гнезд в тычинках покрытосеменных. По геологическому возрасту Williamsoniella и Wielandiella относятся к средней юре.
Несомненные остатки цикадовых встречены были в различных местностях почти во всех отложениях третичного периода; в эоцене Миссисипи, в эоцене Бельгии, в олигоцене и миоцене Чили.
Из голосеменных остаются еще гинкговые и хвойные. Группа гинкговых, в современной флоре представленная лишь одним видом Ginkgo biloba, произошла, по-видимому, от палеозойской ветви семенных папоротников (Pteridospermeae) и обнаруживает точки соприкосновения о палеозойскими и мезозойскими Williamsoniae. Все эти три группы могли иметь схожих предков, но нет никаких данных думать, чтобы они были связаны ближайшими узами родства с современными хвойными. Среди современных деревьев Ginkgo biloba занимает самое изолированное место и является одной из наиболее древних форм. К ископаемым гинкговым относятся, кроме Ginkgoites, еще и род Ваіеrа. Оба эти рода впервые появляются в пермских отложениях и достигают особенного изобилия и разнообразия в триасе, юре в нижнем мелу, где часто можно найти в ископаемом состоянии вместе с листьями их семена, незрелые плодоношения, сережки с тычинками. Ваіеrа, по-видимому, является более древним родом, чем Ginkgo, и апогей ее развития наступил раньше максимума развития последнего. Так, остатки Ваіеrа найдены были в Южной Африке, Южной Австралии и Новой Зеландии.
Группа хвойных Coniferae может быть подразделена на три порядка: 1)Taxales, 2) Araucariales и 3) Pinnies.
Порядок Taxales является наиболее древним среди хвойных; в пользу этого говорит его обширная геологическая история и то, что ареалы распространения его родов в прошлом были гораздо обширнее, чем в настоящее время. В этом порядке можно установить три направления эволюции: 1) семейство Тахасеае, 2) сем. Pudoearpaceae и 3) сем. Phyllocladaceae. Остатки семейства Тахасеае идут вглубь веков вплоть до верхнего триаса (Palaeotaxus). Наличность признаков сходства между Cephalotaxus, Ginkgo, Taxus u Cordaianthus (из кордаитов) говорит в пользу большой древности этого семейства. Семейство Podoearpaceae наших дней является особенно характерным для южного полушария подобно тому, как семейство Тахасеае для северного. Род Podoearpus легко можно назвать господствующим родом среди хвойных южного полушария, так как он найден на всех материках южного полушария, где образует обширные лесные насаждения, которые можно сравнивать с подобными же насаждениями рода Pinus в северных умеренных широтах. Современные Phyllocladaceae ограничены австралийской и новозеландской областями, но оба эти семейства имели обширную геологическую историю. Древнейшим их представителем является р. Palissya из триаса и юры, к которому отнесено около дюжины видов, обильно представленных стерильными ветками в древних слоях мезозоя Европы, Азии и Америки, а также в обоих арктической и антарктической областях.
Podoсarpus содержит около 40 ископаемых видов, которые были широко распространены в течение третичного времени, а некоторые, может быть, и в верхнем меле. Родственный ему род Nageiopsis, представленный в современной флоре секцией Nageia, распространенный от Японии до Ост-Индии и Каледонии, обильно представлен в нижнем меле Северной Америки, а также в вельде Англии и неокоме Азии. О находках его известно из юры Англии и мезозоя Новой Зеландии. Род Phyllocladus проходит через третичные отложения Австралии, а другие, может быть родственные ему роды, вроде Phyllocladites из Шпицбергена и Protophyllocladus из Северной Америки и Азии, представлены в верхнем меле.
Порядок Araucariales в настоящее время представлен двумя родами Dammara (Agatis) в Восточной Индии и Новой Зеландии и Araucaria в Южной Америке и Австралии. Наиболее ранний представитель этого порядка, верхнекарбоновый в пермский род Walchia, имел листья, похожие на игловидные листья современных араукарий; из юрских и меловых отложений из всех частей света описаны несомненные древесины араукарий и чешуи шишек с одной семяпочкой. Листья Dammara и похожие чешуи шишек встречаются также во всех частях света в отложениях верхнего мела. По-видимому, порядок Araucariales характеризовался ареалом в мировом масштабе, так как его несомненные остатки указаны в третичных отложениях Ново-Сибирских островов на Севере Азии, Северной и Южной Америки, Европы, Ост-Индии и на краю Антарктического материка. Лишь Африка не содержит этих ископаемых. Отсутствие их остатков в более поздние времена третичного периода, начиная с плейстоцена, в северном полушарии и их нахождение на Кергуэльских островах и в плейстоцене Фолклендских островов указывает, что Araucariales с этого времени стали сокращать свой ареал распространения.
Последний порядок хвойных — Рinales — содержит 3 семейства: 1) Tаxodiaceae, 2) Cupressaceae и 8) Abietinaceae. Вымершие виды семейства Taxodiaceae во много раз численностью своей превосходят ныне живущие и, очевидно, что семейство прошло уже через кульминационный пункт своего развития; теперь их нет в Европе, Южной Америке и Африке. Геологическая история этого семейства иллюстрирует древность, рост до всемирного господства и последующий затем упадок и распадение на различные типы. Самым древним из известных родов, по-видимому, является Voltzia из пермских и триасовых отложений; в нижнем мелу появляются Sphenolepis и Atrotaxopsis; в верхнем мелу Geinitzia. В конце юры появляется р. Sequoia, остатки которого с того времени в изобилии находят во всех странах за исключением Австралии и Африки. К концу третичного периода ареал ее распространения сильно идет на убыль и в настоящее время ограничивается лишь тихоокеанским побережьем Северной Америки (см. мамонтово дерево).
Семейство Cupressaceae (кипарисовые) появляется в триасе в лице Widringtonites, продолжается в юре в лице Palaeocyparis и Brachyphyllum и сильно развивается в верхнем мелу. Роды Callitris, Cupressus, Chamaecyparis и Libocedrus впервые обнаруживаются в эоцене, как раз в то время, когда многие меловые роды, дожившие до этого времени, успели приобрести широко развернутый ареал.
В семействе Abietineae вымерших родов немного, они очень редки и включают роды Prepinus, Entomolepis и Plutonia из верхнего мела. Среди еще существующих ныне родов р. Pinus обладает многочисленными вымершими видами и может быть прослежен до нижнего мела (Pinites), как и род Cedrue. Рісеа и Abies появляются в верхнем мелу, тогда как р. Tsuga, Pseudotsuga и Larix присущи исключительно третичному периоду и нынешнему времени. Семейство это со времени плиоцена сильно изменилось и широко распространилось в относительно короткий период от олигоцена до наших дней.
Древнейшие ископаемые остатки покрытосеменных (Angiospermae) описаны из различных местонахождений на земном шаре, из отложений, соответствующих гольту (нижний мел), а именно из верхних потомакских слоев Северной Америки, из отложений Vаlаnginien Португалии, из нижнего зеленого песчаника Южной Англии, из слоев Albien Мадагаскара и из отложений Комэ в Гренландии. Кроме того, в 1925 г. опубликована работа Thomas'а о новом порядке Caytoniales из средней юры Англии (нижний оолит Йоркшира). Огромное большинство описанных ископаемых покрытосеменных включено авторами в ныне существующие роды; небольшая часть введена под названием, которое должно отразить принадлежность их к определенному семейству или сходство с отдельным родом (например, Apocynophyllum Ung., Sapotacites Ett, Dryophyllum Deb., Lanropbyllum Göpp. и т. д.); гораздо реже установлены новые роды (Anoectoineria Sap., Crednеria Zenk, Podogonium Heer), или ископаемые растения были описаны под общими обозначениями вроде Antholithes, Carpolithes, Phyllites.
Наибольшую точность в определения ископаемых покрытосеменных допускают цветки, плоды и семена, если они дошли до нас в хорошей сохранности, например, в янтаре, большинство остатков, однако, представлено лишь отпечатками листьев. В морфологическом отношении ископаемые покрытосеменные не обнаруживают никаких существенных различий в сравнении с ныне живущими. Вопрос о происхождении покрытосеменных остается открытым. Если еще для ряда ныне живущих видов ископаемые покрытосеменные позволяют установить ряды предков, то для выведения непосредственных предков для всех покрытосеменных в целом в настоящее время нет достаточных оснований. Палеонтологические находки, в полном согласии с данными систематики растений и географии растений, подтверждают лишь то, что различные семейства покрытосеменных начали развиваться в различных областях земли геологически одновременно. Наиболее древние из покрытосеменных обнаруживают формы, которые с трудом включаются в определенные, ныне существующие систематические единицы; но вскоре обнаруживаются уклоны к различным семействам, особенно из однопокровных, многоплодниковых и однодольных. Лишь в более позднее, меловое время остатки покрытосеменных обнаруживают более несомненные связи с современными растениями, и большинство определимых третичных растений имеет родственников среди современных растений, но они участвовали в сложении тогдашнего растительного покрова в другой пропорции, чем ныне.
В начале третичного периода, во всяком случае, не существовало однородного растительного покрова, который бы покрывал всю землю. Несомненно, существовали различные флористические области, границы которых в течение третичного времени испытали доказуемые сдвиги; равным образом доказано существование во многих местностях вертикального расчленения во флористическом составе. По обе стороны от экватора, как в старом, так и в Новом Свете находились широкие тропические области, границы которых в начале третичного периода простирались в северном полушарии от Южной Англии до Японии и в южном полушарии от Западной Африки до Северной Австралии (область палео- и неотропического элементов флоры по Энглеру). Все остальные части северного полушария (условия южного полушария еще недостаточно точно изучены): полярные страны, Северная Америка, Северная Европа и Северная Англия, составляли одну флористическую область третичной флоры. Относительно определимых остатков этой флоры можно сказать, что их современные родственники живут исключительно в северном полушарии и по характеру своего современного распространения явно могут быть истолкованы как остатки некогда сплошного далеко распространенного растительного покрова (область аркто-третичного элемента флоры по Энглеру). Внутри этих больших поясов уже в древние времена третичного периода существовали области с различными господствующими растительными сообществами, обусловленные различиями в географической широте и тогдашним распределением суши. В смежных областях имело место смешение тропических и аркто-третичных форм, что находит себе выражение в одновременном появлении во флорах эоцена и олигоцена Европы типов растений стран тропических, субтропических и умеренных. В дальнейшем развитии третичного времени мы наталкиваемся на постепенный сдвиг границ этих областей в том смысле, что тропические элементы оказались оттесненными к югу, и в умеренные климаты все более и более стали надвигаться формы растений из полярной области. Аналогичный сдвиг флор в направлении с юга на север доказан и для южного полушария. Тропические и субтропические формы исчезают в плиоцене Центральной Европы; за то на первый план выдвигаются современные растения, среди которых, кроме современных обитателей этих областей, появляются типы северно-американские и восточноазиатские. В течение ледникового периода американские и азиатские формы древней аркто-третичной флоры были уничтожены на материке Европы и лишь в Северной Америке и во внетропических частях Азии (Восточная Азия) сохранились благодаря тому, что орографические условия сохранили для них благоприятные условия существования, вследствие чего мы там и до сих пор можем видеть третичную флору и в цвету и в развитии.
Главнейшая литература по палеоботанике: Seward, А., «Fossil Plants», Vol, I-IV, 1898-1919; Vol I. Ed. II—1925 г. Scotty D., «Studies of Fossil Botany». Edit. III. Vol. I—II, 1920—1923. Scotty D., «Extinct Plants and Problems of Evolution». 1924. Potonie, И. и Gothan, W., «LehrLuch der Palaeobotanik», 1921, Renault, В., «Cours de botanique fossile». Vol. I—IV. 1880—1885. Zeliler, R., «Eléments de Paléobotanique», 1900. Pelourde, «Paleontologie vegetale». Vol. 1. 1914. Залесский, М. Д., «Очерк по вопросу образования угля». 1914, Скотт, Д., «Эволюция растительного мира», 1914.
Л. Кречетович.
| Номер тома | 44 |
| Номер (-а) страницы | 19 |