Мхи
Мхи (Musci), второй класс ряда Bryophyta (мховых, см.). Наиболее развитым является половое поколение (гаметофит); оно-то и представляет собственно мох (рисунок 1). Бесполое же поколение (спорофит) развито весьма слабо, являясь как бы органом или придатком полового поколения и называется, поэтому, спорогонием; оно живет на счет полового поколения и представляет обычно коробочку, сидящую на более или менее длинной ножке. Половое поколение состоит из двух частей: предростка и гаметофора. Предросток (protonema) имеет вид у большинства мхов сильноветвистой, зеленой, весьма тонкой нити (рисунок 2), своими разветвлениями занимающей иногда значительную площадь (до 1 кв. метра) и покрывающей ее в виде зеленой плесени. Ветви нити представляют один ряд клеточек с косыми перегородками. Нить рвется на части, которые продолжают жить дальше, и таким образом мхи могут размножаться. Гаметофор возникает на верхушке боковых коротких ветвей: верхушечная клеточка ветви делится косыми перегородками по трем направлениям; в результате такого деления образуется почка, которая разрастается далее в гаметофор. Предросток бывает двух родов: первичный, возникающий из споры, и вторичный, развивающийся из любой части гаметофора. Гаметофор расчленен на стебель, листья и корневые волоски (ризоиды, рисунок 1). Стебель весьма тонкий и вообще очень незначительной величины, но иногда довольно длинный, например, у кукушкина льна (Polytrichum, рисунок 15) или у тропического мха, Dauwsonia (до метра длиной). Растет стебель при помощи одной верхушечной, трехгранно-пирамидальной клеточки, отделяя сегменты по трем направлениям; из каждого сегмента развивается по листу. Строение стебля довольно простое (рисунок 3): по периферии находится более или менее толстая кора, состоящая из толстостенных склеренхимных клеточек; в середине стебля находится тонкостенная паренхима (сердцевина), а в центре стебля залегает пучок узких, вытянутых в длину волоконец, служащих для проведения питательных веществ; сосудисто-волокнистых пучков у мхов нет, хотя у некоторых из них (Polytrichum, Catharinea и др.) волоконца подразделяются на проводящие воду и растворенные в ней минеральные вещества (ксилемные волоконца) и на проводящие пластические вещества (флоемные волоконца). Типичной кожицы с устьицами на стебле нет. Стебель ветвится. Ветвление обусловливается размещением половых почек, преимущественно почек с архегониями: если они помещаются на верхушке стебля (у акрокарпных мхов), то стебли ветвятся вильчато; если же почки возникают с боку стебля, в пазухе листьев (у плейрокарпных мхов), то ветвление бывает весьма разнообразным. Ветви иногда подразделяются на удлиненные, обычно с редкими листьями, и укороченные, покрытые более густыми листьями. Укороченные ветви бывают либо вегетативные (рисунок 4), либо несут половые органы, а удлиненные иногда развиваются в виде подземных побегов (корневищ). Листья довольно простые: в виде тонких чешуек; нет ни черешка, ни прилистников, ни влагалищ. За исключением жилки, где залегают вытянутые волоконца, лист однослойный, иногда для удержания воды на поверхности листа развиваются тонкие ребра или пластинки (гребни), идущие вдоль листа (у Polytrichum, рисунок 6 и др.). Листья располагаются по спирали и обычно густо налегают друг на друга, так что свободной поверхности стебля не заметно. Часто между листьями развиваются своеобразные выросты в виде зеленых простых или ветвистых нитей или плоских чешуек; это — так называемые парафилии, служащие как для ассимиляции, так и для поглощения и удержания воды. Ризоиды возникают на нижних частях стебля из поверхностных клеточек (рисунок 3); это — длинные ветвистые нити, состоящие из одного ряда клеточек. Ризоиды, находящиеся в земле, бесцветные или бурые, выходящие же на поверхность земли — зеленого цвета; эти ризоиды превращаются во вторичные предростки. Значение ризоидов различно: они служат для поглощения воды и растворенных в ней веществ, а также и для прикрепления гаметофора к почве; это в особенности наблюдается у ксерофитных и эпифитных мхов. Половые органы (архегонии и антеридии) развиваются обыкновенно на укороченных веточках (половых почках, называемых совершенно неправильно цветком). Половая почка появляется либо на верхушке главного стебля (у акрокарпных мхов, рисунки 1, 15 ) или с боку его (у плейрокарпных мхов, рисунки 4, 16, 17). Почка бывает либо обоеполой, когда она содержит архегонии и антеридии, или однополой. Однополые почки размещаются либо на одном и том же гаметофоре (у однодомных мхов, рисунок 5), либо на разных (у двудомных мхов); мужские гаметофоры бывают иногда весьма ничтожной величины (например, у Dicranum, Fissidens и др.), в виде мелких почек, прозябающих между ризоидами женских гаметофоров. Половая почка (рисунок 5) состоит из кроющих листьев, образующих покрывало (или перихетий), и половых органов, разобщенных друг от друга тонкими нитями (парафизами); в мужских почках кроющие листья сильно отличаются и по форме и по окраске от вегетативных листьев, они образуют так называемый перигоний. Архегоний имеет форму бутылочки (рисунок 7) или фляжки, с длинным, узким горлышком (шейкою) и расширенною нижней частью (брюшком); шейка состоит из однослойной стенки и центрального ряда мелких клеточек (канальцевых), которые впоследствии расплываются в слизь; слизь напирает на верхушку шейки и через разрыв ее выходит наружу, так что образуется канал для прохождения сперматозоидов.
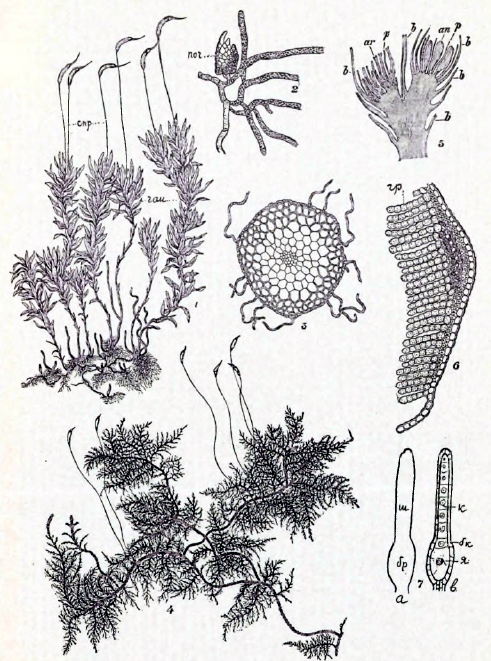
Таблица I.
1 — Catharinea undulata: спр — спорофиты, гам — гаметофиты. 2 — Часть предростка Mnium hornum с почкою (поч.). 3 — Поперечный разрез Bryum roseum. 4 — Thuidium tamariscinum, гаметофит с спорофитами. 5 —Прод. разрез половых почек Phascum cuspidatum с антеридиями (аn) и архегониями (ar), листьями (в) и парафизами (р). 6 — Часть попер. разреза листа Polytrichum comm., гр — гребни. 7 — Архегонии: а — с поверхн., в — в разрезе, ш — шейка, бр — брюшко, к — канальцев. клеточки, бк — брюшн. канальц. клет., я — яйцеклетка.
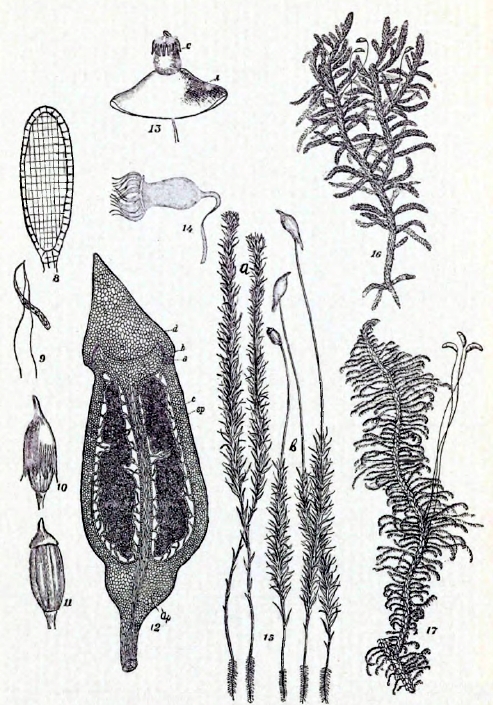
Таблица II.
8 — Прод. разрез антеридия. 9 — Сперматозоиды. 10 — Коробочка Polytrichum com. с колпачком. 11 — То же без колпачка. 12 — Продол. разрез ее: d — крышечка, а — колечко, с — колонка, h — ее расширение, ар — апофизис, sp — споров. мешок. 13 — Коробочка Splachnum luteum, с — окраина, А — апофизис. 14 — Вскрывшаяся коробочка Fissidens adiantoides, с окраиною. 15 — Polytrichum com.: а — гаметофоры с муж. почк. на верхушке, в — гаметофоры с спорофитами. 16 — Hypnum Schreberi: гаметофор с спорофитами. 17 — Ptilium crista castrensis. Гаметофор с спорофитами.
В брюшке помещается крупная яйцеклеточка и мелкая брюшная канальцевая клеточка; последняя перед оплодотворением растворяется, а яйцеклеточка оплодотворяется и дает начало спорофиту. Антеридии имеют форму шарика, эллипсоида или продолговатого или булавовидного тельца (рисунок 8). Состоит антеридий из тонкой однослойной стенки и внутренней многоклеточной (сперматогенной) ткани, развивающей сперматозоиды. Сперматозоид имеет вид продолговатого слегка изогнутого тельца, к переднему тонкому концу которого прикреплены два жгутика (рисунок 9).
Так как сперматозоиды могут двигаться только в воде, то вода имеет существенное значение при оплодотворении. Слизь, выделяемая архегонием, расплывается в окружающей воде и оказывает на сперматозоиды раздражение, привлекая их к архегонию. Обыкновенно в половой почке развивается лишь один спорофит, хотя бы оплодотворилось несколько архегониев. Развитие спорофита происходит следующим образом: оплодотворенная яйцеклеточка, одевшись оболочкой, делится поперечной перегородкой на две клеточки: верхнюю, дающую начало коробочке и ножке спорофита, и нижнюю, развивающую стопу. Возникши в брюшке архегония, спорофит, по мере своего роста, оказывает влияние как на архегоний, так и на соседнюю часть стебля; стенка архегония не только пассивно растягивается, но клеточки ее делятся; у некоторых мхов делятся также и клеточки стебля, соседние с архегонием; благодаря этим делениям, образуется глухая, замкнутая оболочка (epigonium), облекающая молодой спорофит. Разрастаясь дальше, спорофит растягивает эту оболочку, которая под конец рвется у основания архегония кольцевой трещиной. Нижняя часть разорванной оболочки остается при основании спорофита, образуя влагалище, а верхняя часть выносится разрастающимся спорофитом в виде колпачка (рисунки 10, 15b), сидящего под конец на коробочке в виде чепчика или шапочки. Шарообразная, продолговатая, иногда изогнутая коробочка сидит на более или менее длинной ножке (рисунки 1, 4, 15, 16, 17); основание ножки, иногда утолщенное, так называемая стопа, внедряется в стебель и служит для высасывания из него питательных веществ. У некоторых мхов между коробочкой и ножкой развивается еще так называемая шейка (apophysis), которая у Splachnum имеет вид ярко покрашенного зонтика или капюшона (рисунок 13). У некоторых мхов (Sphagnum, Andreaea) ножки нет, но коробочка, тем не менее, приподнимается в воздухе удлиненной верхушкой стебля (ложной ножкой). Внутри коробочки находится бесплодная часть, колонка (рисунок 12), которая у большинства мхов доходит до верхушки коробочки либо (у Sphagnum, Andreaea) не доходит до нее. У многих мхов между спорообразующей тканью, так называемым споровым мешком, и стенкой коробочки замечается воздушная полость, пронизанная ветвистыми тонкими нитями, проходящими от стенки к споровому мешку, а у Polytrichum (рисунок 11) такая же полость находится и между колонкой и споровым мешком. Снаружи коробочка одета типичной кожицей, с устьицами (на шейке). Под кожицей развиваются водяная ткань (иногда в виде одного слоя) и зеленая ассимиляционная паренхима. Внутри коробочки, в споровом мешке, развиваются споры, которые потом высеиваются. По способу вскрытия коробочки мхи могут быть разделены на три группы: 1) клейстокарпных (целоплодных), коробочка которых не вскрывается (Phascum, Ephemerum), стенка такой коробочки выветривается и тогда споры освобождаются; 2) шизокарпных (раздельноплодных), у которых (Andreaea) коробочки вскрываются 4-6 продольными трещинами, и 3) стегокарпных (крышкоплодных) — большинство мхов, у которых коробочка вскрывается крышечкой. Обособление крышечки обусловливается присутствием особого колечка (рисунки 11, 12), находящегося в верхней части коробочки. Колечко состоит из клеточек, разрывающихся при подсыхании коробочки; верхушка коробочки спадает тогда в виде крышечки. Край вскрывшейся коробочки бывает либо гладким, либо снабженным одним или двумя рядами зубчиков (окраиною, рисунки 13, 14). Окраина содействует высеиванию спор, позволяя высеиваться им лишь в сухую погоду и постепенно. Споры микроскопически мелкие, шарообразные; прорастая, они развивают предросток (первичный).
В образовании флоры мхи принимают большое участие, покрывая иногда большие пространства, например, тундры, болота, землю в лесах. В противоположность печеночникам, мхи не приурочены к особо благоприятным (сырым, влажным) местам. Они растут всюду, некоторые в воде (Fontinalis), другие на открытых скалах (Grimmia, Andreaea), на крышах, деревьях; в горах они поднимаются до снеговой границы. По образу жизни мхи делят на 4 группы: 1) ксерофиты — живущие на сухих, большей частью солнечных местах, образуя большей частью плотные подушечки, листья у них снабжены волосками, приспособленными для поглощения влаги из воздуха и атмосферных осадков, таковы виды Tortula, Orthotrichum, Dicranum и др.; 2) мезофиты — живущие на песчаной, каменистой или глинистой почве, насыщенной грунтовой водой, например, Funaria, Bryum, Polytrichum и др.; 3) гигрофиты — растущие на болотах, топях, тундрах, по берегам рек и т.п., образуя глубокие дерновины, например, Sphagnum, Thuidium tamariscinum и др.; 4) гидрофиты — растущие в воде, не образуя дерновин, например, Fontinalis.
Громадный класс мхов, насчитывающий до 12 000 видов, подразделяется на 4 подкласса:
1. Sphagninae. Листья состоят из клеточек двоякого рода: 1) узких, живых, содержащих хлорофилл, ассимилирующих, и 2) более крупных, пустых, мертвых, с кольцевыми и спиральными утолщениями и большей частью с отверстиями. Жилки нет. Колонка не доходит до крышечки; колпачка нет, нет ножки. Коробочка вскрывается крышечкой; окраины нет, предросток пластинчатый. Единственный род Sphagnum, торфяной мох, содержащий до 235 видов, растет густыми дерновинами и подушечками и образует торф. Наиболее обычные виды: S. squarrosum, Girgensohnii cymbifolium и др.
2. Andreaeinae. Листья из одного рода клеточек, с жилкой и без жилки. Колонка не доходит до верхушки коробочки. Есть колпачок; ножки нет. Коробочка вскрывается 4 створками. Один род Andreaea с 105 видами, растущими преимущественно в арктических, подарктических и антарктических зонах, по склонам и камням; наиболее обычен А. petrophila.
3. Archidiinae. Нет ни ножки, ни колонки, ни крышечки. Листья с жилкой. Один род Archidium, до 24 видов, преимущественно в Америке; в Европе только А. alternifolium.
4. Вryinае. Листья с жилкою. Колонка доходит до крышечки. Развиты колпачок, ножка. У большинства видов окраина. К этому подклассу принадлежит большинство мхов, из них наиболее обыкновенны следующие: Саtharinea ur. dalata (рисунок 1), Thuidium tamariscinum (рисунок 4), виды Mnium, Hylocomium splendens, виды Dicranum, Fissidens, Funaria hygrometrica, Hypnum Schreberi (рисунок 16), Ptilium crista castrensis (рисунок 17), Climatium dendroides, Polytrichum commune (рисунок 15) и др. Руководство по М. С. Ростовцева — «Морфология и систематика печеночников и мхов».
С. Ростовцев.
| Номер тома | 29 |
| Номер (-а) страницы | 454 |