Наследственность
Наследственность. Под наследственностью разумеют сохранение и передачу сходного как во внешнем или внутреннем строении, так и в химико-физических особенностях и в жизненных отправлениях организмов. Нередко под наследственностью разумеют какую-то особую силу, что неверно, а также противополагают ее изменчивости (см.), то есть возникновению, появлению несходного, уподобляя различие между ними различию между покоем и движением. Но это сравнение не выдерживает критики, так как понятие наследственности шире и обнимает собой и понятие изменчивости. Удачнее, пожалуй (Тимирязев, Негели), сравнение с принципом или началом инерции, обнимающим оба понятия — покоя и движения («Per inertiam materiae fit ut corpus оmnе vel quiescendi vel movendi difficulter deturbetur». Через инерцию материи происходит, что всякое тело с трудом выводится как из состояния покоя, так и из состояния движения. Newton, «Рrinсiрiа», pag. 2).
Наследственность проявляется как в сохранении неизменного, так и в сохранении изменившегося, только последнее наблюдается далеко не во всех случаях, и вопрос, когда же именно, составляет одну из важнейших современных задач учения о наследственности. Учение это имеет важное значение, теоретическое и практическое, как в отношении к эволюционному учению — дарвинизму (см.), так и в применении к искусству отбора — селекции (см.).
Понятие наследственности обыкновенно приурочивают к явлениям размножения и притом почти исключительно размножения полового у высших растений и животных и в особенности у человека. Это ходячее представление односторонне и не верно. Жизнь отдельного организма представляет смену форм — возрастов, от начального зарождения до смерти. Жизнь вида слагается из преемственной смены поколений. Воззрение на эти две категории явлений зависит от представления об особи, которое далеко не всегда одинаково ясно. Очевидное в применении к человеку и высшим животным, оно далеко не так просто как в применении к сложным животным и в особенности к растениям. В ходячем представлении дерево является особью, размножающейся своими семенами, между тем как на деле это сложный организм; отдельные побеги его — особи последовательных поколений. Люди, удивляющиеся тому, что дети походят на родителей, нимало не удивляются тому, что все листья на дереве одинаковы. В этом случае новые особи происходят из почек, но еще поразительнее и ближе аналогия, когда на срубе старого дерева (например, тополя) отбивается молодая поросль, причем отдельные клеточки образовательной ткани (камбия), в течение веков производившие только определенные элементы коры и древесины, дают начало целым новым особям. На этот раз аналогия еще полнее; особи берут начало не от почек, а прямо от клеточек, как и в процессе полового размножения. Только совершенно забывая значение этих аналогий (которые особенно выдвигал Дарвин), могли создать сохранившиеся до настоящего времени теории наследственности, как пресловутая теория Вейсмана о двух плазмах: зародышевой (Кеimplasma) — вечной и исключительной носительницы наследственности, и телесной (Somatoplasma) — смертной и не имеющей отношения к наследственности. Стоило ботанику (Сидней-Вайнсу) вскоре после возникновения этой теории произнести одно слово — бегония, чтобы разрушить в конец это учение о двух плазмах. У бегонии из надрезов листа, положенного на землю, вырастает целое растение, приносящее цветы и семена, то есть смертная, телесная плазма родит бессмертную носительницу наследственности. Не малую путаницу понятий, по справедливому замечанию Артура Томсона, вносит и распространение на учение о наследственности заимствованного у юристов понятия о наследстве и наследующих. Говорят о переходе к организму того или другого наследства, между тем как в природе наследник и наследство один и тот же объект — наследство это сами наследники или их части.
Ознакомимся с кратким перечнем главнейших категорий фактов, обнимаемых учением о наследственности, а затем уже с так называемыми теориями наследственности. Эти разнообразные категории фактов мы постараемся сгруппировать в порядке их значения, то есть о более общих категориях скажем сначала, а потом перейдем к более частным случаям. Во многих современных трактатах это относительное значение фактов не принимают во внимание, а выдвигают вперед некоторые совершенно частные явления, которым придается несоответственное, а порой даже исключительное значение (как, например, так называемому менделизму; см.).
Прежде всего, следует различать явления наследственности простой и сложной (таблица I, 2, 3). Первая наблюдается при размножении простом, растительном, вегетативном, то есть при превращении в зачаток новой особи части уже существующей взрослой особи. Вторая, то есть сложная, наследственность наблюдается при воспроизведении половом, причем каждая новая особь является наследием двух (явление оплодотворения; см.) организмов, а так как то же повторяется в каждом поколении, то каждый новый организм является наследием всех его предков. Явление это еще более усложняется тем, что у некоторых животных, как это было показано Стенструпом, и у всех растений, начиная со мхов, как это выяснили гениальные обобщения Гофмейстера (см.), полный жизненный цикл каждого организма слагается из правильного неизменного чередования поколений бесполого и полового. Между явлениями размножения полового и бесполого прежде видели коренное различие, но явление партеногенезиса (таблица I, 12) — естественного и отчасти искусственного, — с одной стороны, приближающего явления полового размножения к бесполому, а с другой стороны, возможность искусственного получения растительной помеси путем прививки (таблица I, 11), на что было обращено внимание Дарвином и только в недавнее время немецкими ботаниками, — сблизило эти две основные категории между собой. Наконец, Клебсу удалось показать (см. экспериментальная морфология), что простейшие растения (грибы, водоросли), обладающие обоими способами размножения, можно произвольно превращать в размножающиеся исключительно тем или другим путем.
І. Наследственность простая (таблица I, 2). Наследственность обыкновенно изучают при смене поколений. Периодически повторяющиеся стадии развития каждого поколения начинаются с одной зачаточной клеточки; это одинаково верно как в применении к половому, так и бесполому процессу, но обыкновенно начало нового поколения приурочивают к моменту отделения более или менее сложного или одноклетного тела от производящего его организма. Значение процесса размножения для вида, очевидно, сводится к завладению возможно большей площадью расселения. Простейший способ такого процесса у растений заключается в механическом расчленении взрослого экземпляра на части, будут ли то группы клеточек или целые побеги высших растений, которые так или иначе могут разноситься и превращаться в новые отдельные существа. Лучшим примером этого простейшего случая может служить пресноводное растение Ellodea canadensis, история распространения которого в пресных водах Европы хорошо известна. Появившись в тридцатых годах в Ирландии, оно мало-помалу завоевало Англию, Голландию, Германию, в семидесятых годах пробирается в Оку и т.д. Размножение его идет очень просто: тонкий стебелек его делится вилообразно; отгниванием, разрушением частей ниже развилины выше лежащие части освобождаются, распадаются и разносятся на дальние расстояния. Так как не замечено, чтобы растение это размножалось семенами, то все это один колоссальный экземпляр растения, за несколько лет покрывший своими разветвлениями и отростками всю Европу, а полное сходство его представителей, несмотря на громадное их число и на далекие расстояния, по которым они расселились, служит наглядным примером сохранения сходства путем простой наследственности, почему этот прием так и ценится на практике, когда имеют в виду сохранить какую-нибудь ценную новую садовую или огородную разновидность.
Таблица I
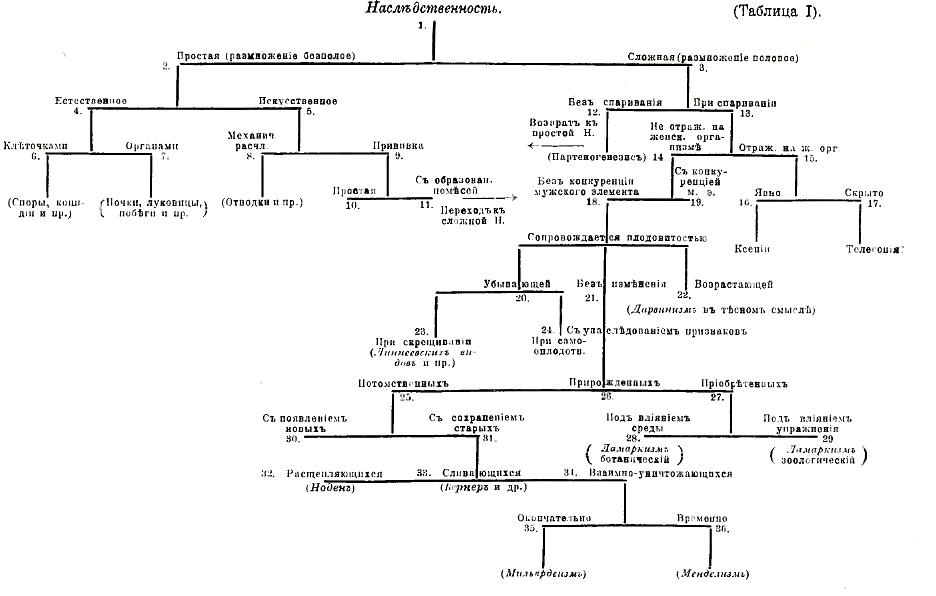
Таблица II

Cytisus Adami - прививочная помесь. C. laburnum и C. purpureus.
Но вегетативное размножение не ограничивается такими простыми случаями; еще чаще части растения, будут ли то отдельные клеточки или более или менее значительные комплексы, образуют своеобразные органы, так или иначе отделяющиеся от растения, — таковы споры или так называемые конидии; таковы различного рода почки, луковички и т.д. Ко всем вегетативным способам применимо то, что сказано о размножении отводками, что при них явления наследственности обнаруживаются с наибольшей очевидностью, то есть сохранение сходного наблюдается с наибольшей полнотой. Эта особенность явления вегетативного размножения была, можно сказать, даже преувеличена: считали, что явления изменчивости при этом способе размножения вовсе не обнаруживаются. Опять Дарвин более, чем кто другой, способствовал рассеянию этого предрассудка и притом с двоякой точки зрения: он собрал обширный материал, доказывающий, что факты изменчивости встречаются далеко не редко и при размножении вегетативном, хотя не так часто, как при половом, потому что в последнем к факторам, действующим в первом случае, примешивается еще новый — совместное влияние двух полов. Согласно представлению, что истинной особью, новым поколением, является образующийся из почки побег, Дарвин указал, что изменчивость должна здесь обнаруживаться в появлении новых свойств у отдельных побегов. Это явление он назвал почковой вариацией, почковой изменчивостью (bud variation), то есть появление, например, на дереве отдельной ветви с темно-красными листьями (как, например, у кровавого бука) или появление на дереве персика ветви с гладкими, а не пушистыми, как это обычно для персика, плодами (нектарины) и т.д. К многочисленным примерам, собранным Дарвином, позднее присоединились еще новые, и дело дошло до того, что самое установление этого особого типа изменчивости стали приписывать другим (например, Коржинскому). Дарвин не ограничился указанием на эту аналогию между двумя категориями наследственности, а указал на другую, еще более неожиданную и глубокую. Он показал, что существуют случаи, когда простая наследственность, свойственная вегетативному размножению, может превращаться в сложную, двойственную, свойственную размножению половому. Словом, он показал, что возможно получение помесей и вегетативным путем, конечно, только искусственно, при содействии человека. Эти случаи встречаются, хотя очень редко, при процессе прививки растений. Идея Дарвина оспаривалась, приводимые им примеры отрицались, все было почти подвергнуто забвению, пока в недавнее время не выступил в ее защиту, подтвердив новыми опытами, немецкий ботаник Винклер. Самый разительный из примеров, приведенных Дарвином, — получение, посредством прививки, помеси между двумя видами Cytisus — Cytisus Іаburnum (часто встречающимся в садах так называемым по-немецки Gold-regen) с желтыми цветами и Cytisus purpureus с красными. Эта совершенно оригинальная по способу ее получения помесь была названа по имени садовода, ее осуществившего в 1829 году, М. Adam — Cytisus Adami. А получилась она таким образом: глазок С. purpureus был привит на стволе С. laburnum, но не принялся, а вскоре на границе дичка и привоя получилось несколько побегов; один из них, высаженный, и дал начало помеси С. Adami. Ее главная особенность заключалась в том, что цветы ее получили окраску, среднюю между С. purpureus и С. laburnum — красноватую. Но этим не ограничилась особенность этой удивительной вегетативной помеси, полученной без полового акта через простое соприкосновение или срастание тканей двух разных видов. Она обнаружила и другое свойство, на которое Дарвин обратил внимание и у настоящих помесей, именно явление, которое он назвал расщеплением (segregation) 1), то есть разъединение у потомства признаков отца и матери между различными ли неделимыми или между частями того же неделимого.
1) Установление этого понятия, проходящего по всей книге Дарвина, Бэтсон потом приписал Менделю.
Оказалось, что С. Adami может на одном деревце приносить троякого рода цветы: и такие, которые свойственны самой помеси, и такие, какие свойственны С. laburnum и С. purpureus, ее предшественникам, так как говорить о родителях здесь было бы неуместно (таблица II, слева цветы С. laburnum, в средине — С. Adami, справа — С. purpureus). На основании этого поразительного и нескольких сходных фактов Дарвин создал понятие прививочных помесей (graft hybrids). «Это образование помесей между видами, в существовании которого я теперь вполне убежден, представляется весьма важным фактом, который рано или поздно изменит воззрения физиологов на половое размножение». Воззрения Дарвина, как уже сказано, были встречены многими учеными крайне скептично.

Фиг. 1. Диаграмма, поясняющая закон потомственной наследности.
Самые факты подверглись сомнению, несмотря на накопление новых, как, например, получение тем же способом, как у С. Adami, прививочного гибрида между двумя родами Сrаtaego — Mespilus помеси боярышника и мушмулы. Только в недавнее время немецкий ботаник Винклер привел новые факты, например, прививочную помесь между томатом и черным пасленом. Впрочем, защитники коренного различия двух способов размножения, (например, Баур) пытаются видеть его в различиях, касающихся строения ядра (см. оплодотворение), но, по Винклеру, в одной его помеси Solanum Darwinianum и ядро представляет среднее строение между двумя видами, послужившими дичком и привоем. Как бы то ни было, в настоящее время не подлежит сомнению, что и при простых вегетативных процессах возможны явления сложной наследственности, хотя бы и не такие полные, как при половом процессе, где происходит более тесное сочетание воспроизводящих элементов.
II. Наследственность сложная (таблица I, 3). Это явление наблюдается при половом процессе размножения, причем в первом же поколении продукты слияния половых элементов должны отражать в себе свойства двух родителей, а принимая во внимание, что организация каждого из них в свою очередь двойственна, получается наследственная зависимость от возрастающего числа предков в той же обратно убывающей прогрессии, что (по Голтону) может быть выражено следующей схемой (фиг. 1). Эта схема, конечно, выражает только теоретически возможную зависимость организации данного существа от организации его предков, в действительности же эта зависимость с первого же шага может представлять уклонения во всевозможных степенях, начиная с полного возврата к бесполому процессу размножения в явлениях так называемого партеногенезиса (таблица I, 14). Явление это заключается в том, что женский половой элемент, яйцеклетка, оказывается способным к образованию нормального организма без участия мужского оплодотворяющего элемента. Всего нагляднее это наблюдается у простейших растений, например, у микроскопического гриба Saprolegnia (фиг. 2), где две совершенно сходные клеточки, образующиеся в женском органе — оогонии, в одном случае вследствие оплодотворения превращаются в ооспору — орган полового размножения, а в другом в партеноспору — орган бесполого размножения.
В некоторых случаях Клебс вызывал явление партеногенезиса искусственно, позднее и зоологи (Лёб, Делаж и др.) получали начальные стадии развития зародыша у морских животных без участия мужского оплодотворяющего элемента, как и Клебс — при действии простых растворов солей. Партеногенезис в сфере явлений полового размножения составляет как бы возвращение к явлениям бесполого размножения точно так же, как возможность вегетативных помесей представляет несомненную аналогию с половым размножением. Известно, что Стенструпом было открыто существование у некоторых групп животных явления так называемого чередования поколений, найденного затем Гофмейстером у всех растений, начиная со мхов. Явление это заключается в том, что полный цикл индивидуальной жизни растения состоит из смены двух поколений, одно из которых размножается бесполым путем, а другое половым; самым типическим, центральным представителем этой коренной особенности растений являются папоротники.
С другой стороны, в связи с простой наследственностью, проявляющейся в вегетативном воспроизведении целых организмов из частей других организмов, можно поставить и явления частичного воспроизведения частей целым организмом, случайно их лишившимся, так называемого явления регенерации, или восстановления органов. В этих случаях, когда на месте утраченного органа восстанавливается новый, совершенно с ним сходный, явления наследственности являются едва ли не в простейшей своей форме. Наблюдаются они и у растений и у животных.
Переходя к изучению сложной наследственности, обнаруживающейся в половом процессе во влиянии оплодотворяющего начала, мы должны, прежде всего, определить сферу действия этого начала. В обыкновенной жизни, особенно в применении к растению, очень часто без всякого удивления готовы признать некоторые проявления этого воздействия оплодотворяющего начала, которые только в самое недавнее время получили объяснение и то еще не во всех случаях. Например, известны различные разновидности маиса, с гладкими или морщинистыми зернами, желтого или синего цвета; еще более известно, что различные сорта яблока бывают различно окрашены и различной формы.
Фиг. 2. Saprolеgniа mixta. А - грибная нить с половыми органами: а — аптеридий, с оплодотворяющим отростком, проникшим в оогоний; о1 — яйцеклетки; о2 — ооспора с образовавшеюся уже оболочкой; ор — яйцеклетки, очевидно без предварительного оплодотворения, превратившиеся в партеноспоры; g — молодой оогоний; В — нить со спорангиями: s1 — зрелый спорангий, проросший сквозь старый опорожненный спорангий; s2 — выхождение зооспор; С — зооспоры и их прорастание: 1 — зооспора в том виде, как она выходит из зооспорангия: 2 — успокоившаяся зооспора; 3 — образование вторичной формы зооспоры; 4 — ее прорастание в нить (по Клебсу).
При опылении одних из этих сортов пыльцой других получаются непосредственные помеси, соединяющие свойства опыляющего сорта со свойствами опыляемого. Ив Делаж рассказывает об одной известной яблоне в Сен-Валери, цветы которой однополы и, следовательно, не производят плодов. Их приходится опылять искусственно пыльцой с других деревьев, и на получающихся непосредственно от этого опыления плодах отражаются свойства сорта, пыльцой которого пользовались. На том же дереве получаются различного сорта яблоки. С первого взгляда, таким образом кажется, что мы имеем здесь дело с обыкновенным получением помесей, но нетрудно убедиться, что это явление совершенно иного порядка, в виду чего для них предложено особое название — ксений (таблица I, 16). В семени будущему поколению соответствует только зародыш, несомненно, происходящий через слияние двух половых элементов. Уже и в семени имеются части, составляющие непосредственно части исключительно материнского организма, например, так называемый белок и оболочки семени (как у маиса); это еще очевиднее в применении к плоду, представляющему изменившуюся после оплодотворения завязь женского цветка. Таким образом, влияние оплодотворяющего начала может отражаться не только на потомстве, но и на организме матери. Явление это долго представлялось загадочным, пока Гиньяру и Навашину не удалось показать, что у растений в зародышевом мешке происходят одновременно два процесса: процесс оплодотворения яйцеклетки, дающий начало зародышу, и другой процесс — оплодотворения (слияние двух других ядер), вызывающий изменения в остальных частях будущего семени (см. оплодотворение). Менее ясны влияния оплодотворяющего начала на еще более отдаленные части материнского растения. Некоторые ученые даже прямо отрицают их, но для этого, в виду ясно исследованных случаев (например, опыты Гильдебранда), едва ли есть повод, особенно со времени открытия гормонов (см.) — веществ, распространяющихся в организмах и вызывающих органические изменения на далеких расстояниях (каково у животных, например, влияние гормонов, выделяемых зародышем, на развитие молочных желез организма матери). В явлении ксений усматривается непосредственное влияние оплодотворяющего начала на организм матери, но рядом с ними наблюдалась и другая категория явлений, где о таком воздействии заключают не по непосредственному влиянию на организм матери, а по влиянию при посредстве его на последующее ее потомство уже от другого отца. Типическим примером этого явления, получившего название телегонии (таблица I, 17), служит игравший большую роль в истории этого вопроса случай с «кобылой лорда Мортона». Кобыла, однажды спаренная с кваггой, при последующих случках с жеребцом дала потомство с явными признаками квагги, полосами на ногах и т.д. Этот случай долго приводили в литературе как несомненный; придавал ему веру и Дарвин. Наконец, в 1899 году Юарт (Ewart) документальными изысканиями убедился в сомнительности этого знаменитого случая, а своими собственными тщательными опытами (известными в литературе под названием Пениквикских опытов) над кобылой и зеброй показал несуществование этого явления телегонии. Таково фактическое положение этого вопроса, хотя и здесь только что упомянутые воздействия гормонов могли бы быть выдвинуты против предъявляемых теоретических возражений.
Переходим, наконец, к самой существенной части явлений наследственности, сводящейся к взаимодействию между половыми элементами, и остановимся на сопровождающих оплодотворение последствиях: на различных степенях плодородия, на различных степенях наследуемости признаков, на различных способах их сочетания и т.д.
По отношению к эволюции организмов, результат, прежде всего, должен зависеть от того, какой оплодотворяющий элемент будет иметь более шансов на достижение к оплодотворяемому элементу. В большинстве случаев, как, например, у животных, это уже относится к области явлений полового отбора, но в некоторых случаях эта борьба не так очевидна, не так ясна. Так, например, у высших растений на поверхность одного рыльца может попадать пыльца различных растений, но результат оплодотворения не зависит от случая, а всегда наблюдается, что между конкурентами находятся обладающие каким-то преимуществом перед своими соперниками (явление, названное Дарвином преобладанием, осиливанием, prepotency; таблица I, 19). Но отличие этого случая от полового отбора, конечно, не существенное, так как крупинки пыльцы соответствуют, в сущности, целым мужским организмам. Занимающие нас явления наследственности начинаются только с момента взаимодействия половых элементов. С упомянутым явлением prepotency необходимо, однако, считаться при опытах скрещивания, чтобы не смешать осиливающие организмы (пыльцу), отчего зависит самая возможность, оплодотворения с осиливающими отдельными признаками, отчего зависит только качественный результат оплодотворения.
Главным результатом оплодотворения, с эволюционной точки зрения, является, конечно, большая или меньшая успешность его, то есть степень плодовитости в первом же или в последующих поколениях. Если отправляться от какой-нибудь средней обычной плодовитости между сходными представителями того же вида, которую принять за нормальную (таблица I, 21), то можно встретить отступления как в сторону убывающей плодовитости вплоть до полного бесплодия (таблица 1, 20), так и в сторону возрастающей плодовитости (таблица I, 22). Здесь, по-видимому, наблюдаются две закономерности: во-первых, уменьшение плодовитости до полного бесплодия, когда различие форм приближается к границам «хороших», или «Линнеевских» видов (таблица I, 23), а также в некоторых случаях самооплодотворения (таблица I, 24). С другой стороны, исследованиями самого Дарвина обнаружена и прямо противоположная закономерность — возрастание плодородия и более могучей организации (constitutional vigour) у помесей между несколько отличающимися между собой (по происхождению или местонахождению) представителями одного вида в сравнении с результатами процессов самооплодотворения. Этот последний закон давно должен был бы получить название закона Дарвина, или дарвинизма (таблица I, 22) в тесном смысле слова (в отличие от его основной теории — дарвинизма).
Он является объяснением для роли бесчисленных, наблюдаемых в природе приспособлений, клонящихся к обеспечению перекрестного оплодотворения, а может быть, и ключом для объяснения самого возникновения полового процесса. С ним также необходимо считаться при всех опытах получения помесей, на которых основываются все новейшие соображения о наследственности. Так, например, он дает ключ для объяснения кажущихся непонятными результатов помеси двух форм — мелкой и крупной, когда, вместо средней между ними, получается более крупная, чем крупная. Такие результаты получались, например, у Менделя и совсем необъяснимы, исходя из его правила. Этот же закон Дарвина объясняет и поразительные результаты в опытах Бурбанка, когда при скрещивании двух форм грецкого ореха он получил не среднюю из двух, а значительно более могучую, более крупную, чем крупная, и скороспелую новую породу. Подтверждению этого закона Дарвин посвятил несколько лет; результаты опытов составляют целый том («Cross and Selffertilisation of Plants», 1876), и тем не менее в больших современных трактатах о наследственности о нем или вовсе не упоминается, или результаты приводятся с указанием на их важное значение, но тогда они приписываются другим ученым или приводятся анонимно, (например, у Иоста в его курсе физиологии растений).
За этими общими проявлениями сложной наследственности переходим уже к более частным, относящимся к наследованию отдельных признаков, к изучению, какие из них и в какой степени передаются. Во многих современных трактатах можно встретить утверждение, будто только со времени Менделя стали говорить о наследственности отдельных признаков, а прежде будто бы имели в виду огулом всю совокупность признаков того или другого родителя. Между тем в своей книге с начала и до конца Дарвин говорит о наследственности отдельных признаков (inheritance of characters), особенно подчеркивая тот факт, что самые ничтожные из них могут передаваться в отдельности.
Прежде всего, обращают внимание на две основные категории признаков: на признаки прирожденные и признаки приобретенные.
Под прирожденными очень часто смешивают как те признаки, которыми обладали родители и предки рассматриваемой формы, так и те, которые приобретены ею самой в период эмбриональной жизни; под приобретенными — те, которые возникли вновь при условиях существования уже развитого организма, но, строго говоря, сюда правильнее следовало бы относить и все изменения, относящиеся к периоду эмбрионального развития, так как известно, что полным сходством обладают исключительно формы, не только имеющие одинаковую наследственность, но и одинаковые условия эмбрионального развития, то есть близнецы. Факт этот доказывает, что влияние условий существования не ограничивается периодом после рождения, но определяется и периодом от момента оплодотворения и до рождения. Так как влияния внешних условий тем глубже, чем ранее они начинают действовать (см. изменчивость), то можно ожидать, что и передача их путем наследственности будет более обеспечена. Может быть, было бы правильнее различать троякие признаки: потомственные (таблица I, 25), общие с родителями и более отдаленными предками; прирожденные (таблица I, 26) и признаки приобретенные (таблица I, 27), считая последние две категории за сходные по происхождению, но различные по глубине воздействия внешних условий, а потому и различные по степени их унаследования. Едва ли не наибольшее внимание сосредоточено в последнее время на последней из трех категорий. Едва ли какой вопрос в области наследственности подал повод к такой оживленной, порой ожесточенной, полемике, как вопрос о наследственности приобретенных признаков. Одни категорически его отрицают (как, например, Вейсман), другие основательно возражают, что без его допущения немыслимо никакое эволюционное учение (Спенсер). Наконец, немалую смуту внесли в обсуждение этого вопроса так называемые неоламаркисты, путающие две различные точки зрения, высказанные Ламарком: по отношению к растениям и к животным (см. Ламарк). Поэтому, прежде всего, этот вопрос о наследственности приобретенных свойств нужно разделять на два: на наследование изменений, вызванных действием внешних факторов (среды, milieux ambiant старых французских биологов), и наследование изменений, вызванных деятельностью самого организма, главным образом упражнением (или неупражнением) органов. Эту последнюю категорию Плат Болл, автор прекрасного критического этюда по этому вопросу, называет, в отличие от первой, use inheritance; новейшие немецкие авторы называют ее функциональной наследственностью. Известно, что Ламарк придавал выдающееся значение именно этому, по его мнению, важнейшему источнику изменчивости, видя в нем сознательное участие воли самого существа, чем объяснялось бы образование форм, приспособленных к их условиям существования. Дарвин вначале отрицал значение этого Ламарковского фактора, но позднее принимал его более в отрицательном смысле, то есть в смысле атрофирования, вырождения органа вследствие отсутствия его упражнения, как, например, крыльев у нелетающих птиц. Основная идея Ламарка выражается в известной французской поговорке: «à force dе forger on devient forgeron», но вопрос, родятся ли дети кузнецов с готовыми мускулами кузнеца, на опыте неизменно разрешается в отрицательном смысле. Плат Болл приводит даже пример тому, что существование такой наследственности явилось бы прямо роковым. Известно, что развитие мышц атлетов, учителей фехтования и т.д. нередко ведет к параллельному перерождению мышц сердца порой со смертельным исходом. При наследственном накоплении этой особенности, орган этот перестал бы служить для своего отправления. По отношению ко второй и более важной категории приобретенных свойств — тех, которые являются результатом воздействия внешних условий (действующих физиологически или патологически), как уже сказано, мнения расходятся, и прежде всего здесь, вероятно, следует допустить целую градацию явлений. Категорически отрицающие возможность наследственной передачи какого-нибудь приобретенного в течение явной (то есть не эмбриональной) жизни признака иногда опираются на опыты, состоящие в грубом механическом повреждении: так, например, Вейсман обрубал хвосты мышам и не наблюдал у них куцего потомства. Бездоказательность такого опыта очевидна. В применении к отдельному организму нужно, очевидно, руководиться тем же представлением, как и в применении к родственной преемственности, выражаемой родословным деревом. Изменения в организации дяди не могут влиять на организацию племянника, точно так же и клеточки ткани хвоста не представляют предков, воспроизводящих тех клеточек, из которых разовьется потомство мыши с обрубленным хвостом. Строго говоря, можно защищать и обратное, то есть не требовать непременно родословной преемственной связи между элементом, воспринявшим действие внешнего фактора, и элементом, передающим его последующему поколению. То, что сказано было выше о действии гормонов, может найти применение и здесь. Иное дело воздействия, отражающиеся на всей организации потерпевшего организма, физиологические или патологические, как, например, в классических опытах Браун-Секара над искусственно вызванной падучей болезнью морских свинок. Они прямо или косвенно влияют и на воспроизводительную систему, то есть на начальные элементы будущего организма. Понятно, что проследить непосредственную, непрерывную цепь причинной связи между воздействием среды на предшествующее поколение и его передачей следующему поколению, в чем и заключается истинная задача исследования наследственной передачи приобретенных признаков, почти невозможно и до сих пор не удавалось. Остаются только попытки эмпирического удостоверения, что такие-то изменения, вызываемые искусственно, передаются, другие не передаются. Разрешение вполне этой задачи входит в область экспериментальной морфологии (см.), но пока число достоверных случаев прочного унаследования изменений, вызванных искусственно, очень ограничено. Главное затруднение заключается, вероятно, в том, что здесь важную роль играет фактор — время. Быть может, воздействие в течение одного поколения не оставляет еще прочного следа, между тем как воздействие в течение нескольких поколений оставило бы по себе прочный наследственный след. Многие специалисты (Бэли, Вильморен, Костантен) высказывали эту мысль в такой удачной форме: говорят о наследственности приобретенных свойств, но сама наследственность — не является ли она приобретенным свойством? Одним из убедительных примеров наследственной передачи внешнего воздействия служит махровость цветов, так как, с одной стороны, это явление вызывается искусственно, а с другой, оно, несомненно, наследственное. В области простейших растений указывают на опыты Ерреры над одним плесневым грибком, у которого ему удалось показать наследственную передачу некоторых искусственно вызванных осмотических свойств. Мечников указывает на общий факт, что некоторые физиологические разновидности бактерий, возникающие в ненормальных для них условиях, наследственно сохраняются и при возвращении последующих поколений в нормальные условия. Известному американскому зоологу Тоуэру удавалось вызывать под влиянием высокой температуры и т.д. изменения в окраске колорадского жука и показать, что эти изменения наследственны. Разрешение этого вопроса строго экспериментальным путем, как уже сказано, — задача будущего, задача экспериментальной морфологии, но некоторые ученые полагают, что и путем простого наблюдения можно придти к выводу о наследственности приобретенных признаков, указывая, что этот путь, если не так строг, то зато убеждает многочисленностью фактов, говорящих в его пользу. Таковы воззрения Костантена и особенно Генсло. В книге, так и озаглавленной: «The inheritance of acquiered characters» («Унаследование приобретенных признаков»), Генсло, опираясь на многочисленные категории «бесчисленных», как он выражается, фактов, развивает следующую аргументацию. На различных точках земного шара представители самых различных групп растительного царства обнаруживают поразительные черты сходственного строения, если только местные условия их обитания сходны (сухость и влажность, температура, воздушная или водная среда, отношения к другим растениям, то есть паразитные и вьющиеся растения, и т.д.). Это общий вывод современной физиологической географии растений. Сходства эти не только внешние, но и внутренние и притом охватывают признаки, которым придавали значение не только для установления разновидностей, но нередко видов, родов и т.д. до самых обширных систематических групп (например, однодольных и двудольных). Присутствие одинаковых форм неизменно при одинаковых условиях, принимая во внимание громадное число фактов, уже оправдывает с точки зрения вероятности заключение, что мы имеем здесь дело не со случайным совпадением, а с причинной связью, то есть что эти формы являются результатом воздействия этих условий. Но эта высокая степень вероятности превращается почти в полную достоверность, если на небольшом числе случаев удается на опыте показать, что именно такие формы действительно вызываются именно такими условиями. Наконец, хотя в большей части случаев не зарождается даже сомнения, что эти признаки, связанные с условиями обитания, наследственны, — это положение можно доказать теми случаями, когда организмы, будучи перенесены в другую среду, еще некоторое время сохраняют свою прежнюю форму уже не в силу воздействия среды, более не существующего, а в силу наследственности. Таким образом, по Генсло, имея в виду возможно большее число растений в возможно разнообразных географических условиях, мы невольно выносим общее заключение, что целый ряд их особенностей вызван продолжительным влиянием условий их существования, и в силу этой продолжительности особенности эти так закрепляются, что по инерции некоторое время сохраняются даже в отсутствии вызвавшего их влияния (в чем и выражается наследственность), пока не подчинятся действию новой среды. По мнению Генсло, наблюдение бесчисленных примеров представляемого растительным миром всего земного шара отношения к среде, освещенное несколькими физиологическими опытами именно такого непосредственного воздействия среды на растительные формы, служит одним сплошным аргументом в пользу разрешения в положительном смысле вопроса о том, наследуются ли приобретенные признаки. Это, можно сказать, основное положение эволюционного учения, о котором Спенсер метко выразился: «или существует наследственность приобретенных признаков, или не существует эволюции». Отсюда понятно, что такой противник какого бы то ни было эволюционного учения, как Бэтсон, желая от него окончательно отделаться, в самое недавнее время предложил воззрение, представляющее в буквальном смысле возвращение к старому изречению: «ничто не ново под луною». Он утверждает (1914 г.), что никакой изменчивости не существует, ничего нового не возникает, происходят только, в силу скрещивания, перетасовка, новые сочетания от века существовавших свойств, и то, что ныне кажется простым, в действительности сложно, так как в кажущемся простом находится готовым уже все то, что нам позднее представляется сложным. Ограничиваясь сказанным по отношению к вопросу, составляющему главную задачу будущего развития экспериментальной морфологии, этой основы эволюционного учения, переходим к рассмотрению тех проявлений наследственности, в которых участие наследственности уже не подвергается сомнению (наследственности потомственной и прирожденной). Весь интерес на этот раз сосредоточен на том, как сочетаются между собой наследуемые свойства, какие закономерности наблюдаются по отношению к участию двух сторон, двух родителей, определяющих свойство сложного потомства.
Таблица III.

A. Medicago sativa. B. Medicago falcate. C. Помесь A. и B. – Medicago media.
С акварели А. Н. Строганова.
Здесь снова, прежде всего, следует различать два случая: во-первых, сочетание в потомстве разнородных частей организации и, во-вторых, сочетание различных свойств однородных частей или, выражаясь вообще, одинаковых особенностей обоих родителей. В первом случае собственно нет столкновения, взаимодействия или противодействия двух наследственностей, во втором оно, так или иначе, проявляется, и здесь-то и выступают различные закономерности, сосредоточивающие почти все внимание новейших исследователей. Первое явление — совмещение различных особенностей, не находящихся в прямом антагонизме, являющееся едва ли не главным источником эволюции в смысле совмещения, накопления полезных свойств — представляется главным условием отбора как искусственного, так и естественного. Успех этих комбинаций в последнее время нередко приписывают применению учения Менделя (например, опыты Биффена над сочетанием плодовитости одних пород пшеницы с устойчивостью другой от заразных болезней). Но прием этот практиковался задолго до появления менделизма и без его применения; так, Вильморен получил помесь пшеницы Dattel, соединившую свойства соломины одной со свойством зерна другой породы; сюда же относится вся деятельность Бурбанка (см.), с менделизмом не имеющая ничего общего.
Можно сказать, что почти весь интерес современных исследований сосредоточивается на втором случае, на изучении результатов скрещивания между родителями, представляющими различия в однородных частях или, выражаясь вообще, в однородных особенностях. Только в этом случае и возможно наблюдать результат столкновения двух наследственностей. Эта группа явлений, в свою очередь, может представлять два случая. Первый из них — когда получаемая помесь несет только следы своего двойственного происхождения, и второй — когда, помимо особенностей, присущих родителям, проявляются, по-видимому, совершенно новые черты (таблица I, 30, 31). Этот второй случай (таблица I, 31) объясняется иногда очень просто, — взаимодействием между явным признаком одного организма и скрытым признаком другого. Корренс приводит такой простой убедительный пример: существуют две разновидности садовой льнянки: одна розовая, другая белая; можно ожидать, что средняя между ними будет бледно-розовая, а получается сине-фиолетовая, то есть, по-видимому, нечто совершенно новое. Дело в том, что белые цветы, кроме отсутствия красной окраски, отличаются еще присутствием щелочной реакции, а щелочь, как это известно из реакций лакмуса, окрашивает пигмент розовых цветов в фиолетовый, что и наблюдается у помеси. Таким образом, получение совершенно новых признаков иногда объясняется очень просто, но, конечно, оно может зависеть и от более сложного взаимодействия смешивающихся организаций, требующего обстоятельных исследований.
Переходим, наконец, к подробно изученным случаям образования помесей, в которых ясно обнаруживаются сложные влияния родителей, и познакомимся с обнаруженными при их изучении закономерностями.
Здесь снова приходится отметить три случая (таблица I, 32, 33, 34): наследственность смешанную, или расчлененную (disjointe по Нодену), наследственность слитную и наследственность взаимно-исключающуюся, то есть признаки родителей: или только перетасовываются, то есть остаются разделенными в пространстве, представляя явления так называемой мозаичности; или сливаются между собой, как смешивающиеся жидкости, давая начало признакам, средним между двумя: или, наконец, взаимно исключаются, то есть в помесях признаки одного родителя вытесняют признаки другого.
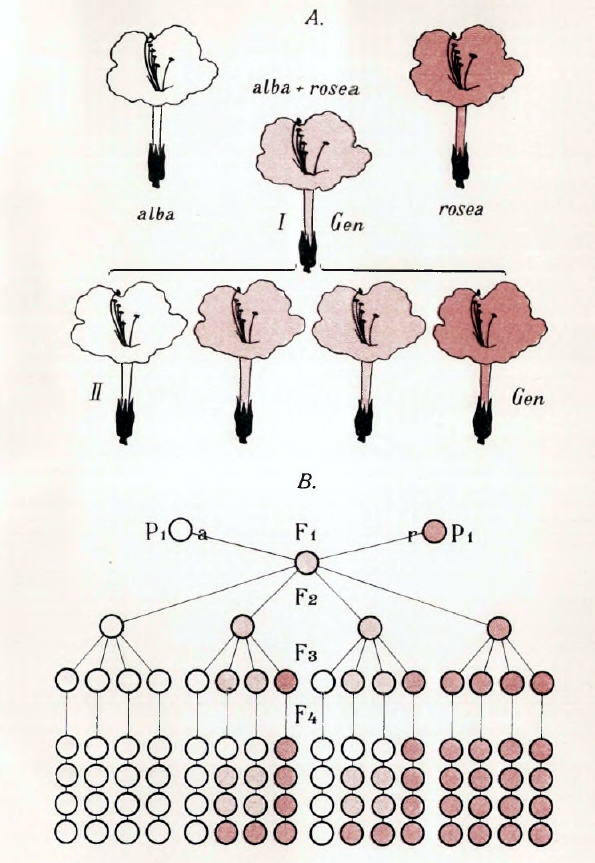
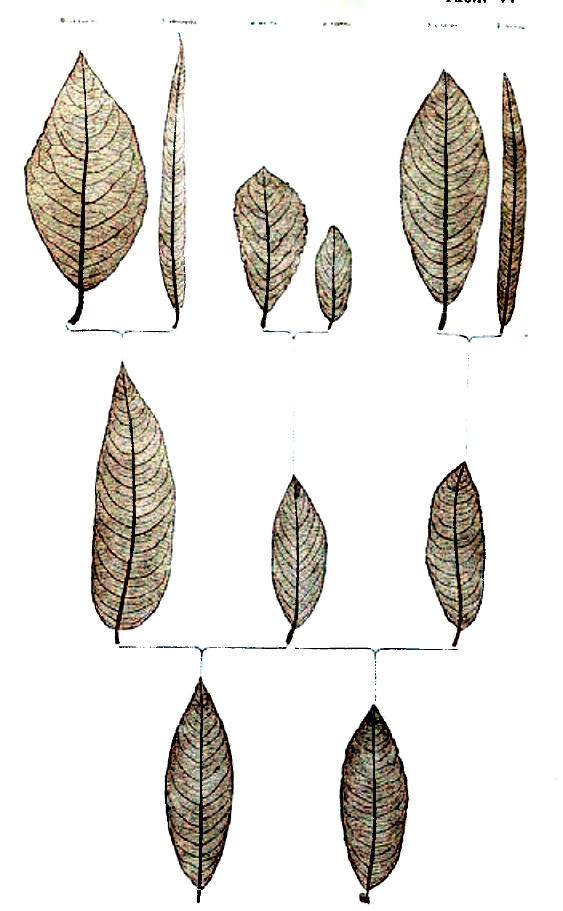
Наследственность смешанная представляет смешение признаков двух скрещивающихся форм, так что они оказываются разделенными в пространстве: одни участки организма помеси воспроизводят свойства одного родителя, другие — другого, как это наблюдается при смешении двух окрасок двух мастей в одну пятнистую или пегую. Таковы, например, полосатые лепестки цветов или пегие животные. Этот случай интересен как по очевидности совмещения признаков двух родителей, так и по тем теоретическим выводам, которые сделал из этих фактов, как увидим ниже, для общего учения о наследственности ботаник Ноден, назвавший эти случаи hybridation disjointe — скрещиванием расчлененным. Наследственность слитная. Этот второй случай наиболее простой, наиболее естественный, наиболее понятный и давно подмеченный как в природе, так и в практике садовода и скотовода. Некоторые ученые, как, например, Кёрнер, видят в нем главный источник образования новых форм, новых видов и ссылаются на многочисленные примеры. Остановимся на одном из них (таблица III); это помесь двух видов люцерны: Medicago sativa — посевной и Меdicago falcata — желтой, получившая название Medicago media, люцерна средняя, или промежуточная. Этот средний характер всего нагляднее обнаруживается в форме плода и в окраске цветов. У посевной бобы, закрученные пробочником с тесно сближенными оборотами спирали; у желтой бобы серповидные; у средней бобы представляют 11/2-2 разодвинутых оборота спирали. Цветы у первой сине-фиолетовые, у второй — желтые, у средней — грязно-зеленые, окраски совершенно необычной для цветов. Эта окраска является результатом смешения сине-фиолетового клеточного сока первой с желтыми зернами второй. Остановимся еще на одном примере образования промежуточной помеси и притом на таком, который выяснит нам тот основной числовой закон, который вытекает из новейших многочисленных исследований в этой области. Это, обстоятельно изученная Корренсом, помесь двух разновидностей Mirabilis Jalappa — розовой и белой (таблица IV А). Помесь между ними roseaXalba имеет цветы розовые, но более бледные, чем у rosea, то есть окраски средней между обоими родителями. И результаты скрещивания могут не ограничиваться одним поколением. Скрещенные формы могут, в свою очередь, подвергнуться скрещиванию и совместить в одной организации признаки уже не двух, а четырех и большего числа организмов. Это всего нагляднее выступает из опытов, произведенных в 1865 году Вихурой. Он скрещивал различные виды ив, обращая внимание главным образом на форму листа. Три полученные помеси, давшие три средние формы, скрещивались между собой и давали помесь второго порядка, среднюю между четырьмя предками. Вот результаты одного такого опыта (таблица V). Возвращаемся к помеси Mirabilis Jalappa roseaXalba, r + a или а + r, что обыкновенно безразлично, хотя есть случаи, когда и не безразлично. Пример этого последнего дают два другие вида той же — Mirabilis Jalappa и longiflora; оказывается, что одно скрещивание — первой пыльцой второй — вполне плодовито, а обратное бесплодно, и объясняется это очень просто; цветневые трубочки первой слишком коротки для длинного столбика второй; оплодотворение механически невозможно. Возвращаемся к помеси розовой и белой разновидности (таблица IV А). Если цветы, r + а опылять собственной пыльцой, то получается нечто более сложное. В первом скрещивании r и а (таблица IV, АІ) организация обоих родителей была простая, а во втором случае организация каждого родителя будет уже двойственная, и при сочетании этих двух двойственных организмов возможны уже четыре случая аха, ахr, rха, rхr; первая из них будет чистая, а вторая и третья — помесь обеих, а четвертая чистая r (таблица IV АII). Любопытно, что в том же 1865 году два человека одновременно остановились на мысли, почему получается именно такой результат; но один более остановился на объяснении, почему это происходит, а другой — на том, каков должен быть численный результат такого самоопыления у помеси, совмещающей свойства двух различных разновидностей. Ноден задумался над тем, как могут от скрещивания сложной формы получиться обратно простые. Очевидно, соединяющиеся в помеси качества организмов отца и матери могут разъединяться, расчленяться (se disjoindre), а при вторичном оплодотворении сочетаться и сходные со сходными и несходные между собой. Но как представить себе фактически это расчленение.
Таблица IV.
Помеси двух разновидностей Mirabilis Jalappa — alba и rosea.
А. Цветы двух родителей и двух поколений помеси.
В. Статистическая схема двух родителей и четырех поколений помеси (По Корренсу).
Реальный ключ к этому, по его мнению, дает первая из перечисленных нами категорий образования помесей, которую он и назвал помесью расчлененной — hybridation disjointe (таблица I, 32), состоящей в том, что признаки отца и матери не сливаются в один общий средний признак, а завладевают различными участками организма помеси. Пример этого нами изложен у Cytisus Adami, где рядом с цветами средней формы появляются соцветия с чистыми цветами формы отца или матери. Но это расщепление (segregation Дарвина) может, конечно, идти еще глубже; оно может охватывать отдельные участки ткани; так, например, при скрещивании двух видов той же Mirabilis — Mirabilis Jalappa и Mirabilis longiflora, белой и красной, эти пигменты не сливаются в светло-розовый, а получаются лепестки пестрые с полосами белыми и красными. В виду этого факта, рассуждает Ноден, сделаем еще один шаг и предположим, что качества могут расщепляться между отдельными клеточками пыльцы или яйцеклетками, и тогда станет понятна возможность одновременного образования формы в отца, в мать или средних между ними. В пользу этого блестящего объяснения, Мильярде привел позднее (1894 г.) следующее изящное наблюдение. Пример Нодена, остановившегося на расчленении признаков по отдельным участкам тканей, Мильярде действительно доводит до отдельных клеточек. Он изучал помеси различных видов винограда и нашел, что устьица кожицы листа представляют у них очень характеристичные формы, например, у Vitis labrusca и Vitis aestivalis, фиг. 3. Если получить помеси между ними, то можно найти листья с устьицами, как у отца и как у матери, и промежуточные между ними, фиг. 3. Но аппарат устьиц происходит из одной клетки, следовательно, в кожице листа встречаются клетки типа отца и клетки типа матери и клетки типа помеси, отстоящие одна от другой на расстоянии каких-нибудь десятых долей миллиметра. Это прямое наблюдение Мильярде делает вполне реальной гипотезу Нодена, то есть что при таких же условиях и отдельные клеточки пыльцы и еще легче отдельные яйцеклетки могут иметь обособленный характер отца или матери или продукта скрещивания между ними. Далее эти расчлененные крупинки пыльцы или яйцеклетки могут скрещиваться с нерасщепленными, и таким образом получатся помеси квартероны и т.д. Все свои рассуждения Ноден заканчивает выводом: «окажутся возможными всякие сочетания, и управлять ими будет случай».
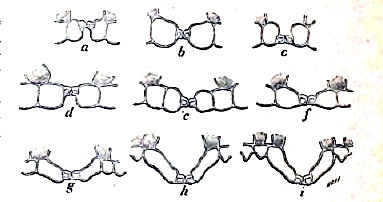
Фиг. 3: a, b, c, - устьица Vitis acstivalis: d, e, f, g, h – устьица помеси V. e. и V. l.; i – устьица Vitis labrucsa (по Millardet).
К тому же выводу и в том же 1865 году пришел Мендель, но если Ноден привел более реальные обоснования для возникновения самого явления расчленения (hybridation disjointe Нодена, segregation Дарвина), то Мендель сделал из этого заключения дальнейший числовой вывод, — недаром учился он математике в венском университете! Если этим явлением управляет «случай», то — рассуждал он — при надлежащем изучении должны обнаружиться и те законы, которые управляют случаем, то есть помеси и чистые формы должны представлять те численные отношения, на которые указывает теория вероятностей. В простейшем случае, когда дело касается двух признаков, возможные случаи выразятся так:
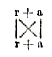
то есть rr + rа + аr + аа, как и оказывается на деле, по одной чистой форме r и а и две смешанных, между собой тождественных (таблица ІV В). К сожалению, как будет сказано в своем месте ниже, Менделю подвернулся очень неудачный пример (горох желтый и зеленый), затемнивший основной закон совершенно частным из него исключением, которое его слепые поклонники возвели в общий закон, стараясь насильственно подвести под него все противоречащие случаи. Закон Нодена-Менделя, по которому потомство помеси при ее самооплодотворении дает начало не только средним формам, но воспроизводит и чистые формы родителей, имеет, очевидно, громадное значение для эволюции организмов, так как показывает, что скрещивание вновь появившихся форм не грозит им уничтожением, а представляет для естественного отбора широкий выбор между чистыми и смешанными формами, чем устраняется то возражение против дарвинизма (в Англии высказанное Флиммингом Дженкинсом, у нас повторенное Данилевским), которое и сам Дарвин признавал самым опасным для его теории.
Наследственность взаимно исключающаяся. Наконец, третий общий случай образования помесей заключается в том, что признаки скрещивающихся организмов оказываются не совместимыми, как в первых двух, а один из признаков вытесняет, осиливает другой. Этот общий случай, в свою очередь, распадается на два частных случая. Первый из них заключается в том, что вытесняющий признак окончательно осиливает другой и сохраняется в потомстве, будто никакого скрещивания не происходило. Этот случай тщательно исследован французским ботаником Мильярде над несколькими видами земляники и назван им hybridation sans croisement — гибридация без скрещивания. Эту главу учения о наследственности с таким же правом можно назвать мильярдеизмом, с каким главу, трактующую о следующем втором случае, называют менделизмом. Этот второй случай, составляющий содержание того, что собственно лишь имеет право называться менделизмом (см.), заключается в следующем. Из двух скрещивающихся форм признак одной оказывается осиливающим, доминирующим (другой признак называется в отличие рецессивным), и все первое поколение обладает исключительно им (скрещивание желтого гороха с зеленым дает первое поколение исключительно желтое, см. Мендель). От явлений мильярдеизма явления менделизма начинают отличаться только со второго поколения, получаемого путем самоопыления. Это второе, как и последующие поколения, являются смешанными: горох будет и желтый и зеленый — и притом в установленном Менделем числовом отношении — на три желтых один зеленый. Как указано выше, общее объяснение закона, управляющего образованием помесей, предложено Менделем (и одновременно Ноденом в том же 1865 году), но только общий случай (а + ab + ba + b) в силу доминирования одного признака превратился в За + b, то есть три желтых и один зеленый. Этот-то совершенно исключительный частный случай, только затемняющий смысл основного закона, фанатические поклонники Менделя желают возвести в основной закон наследственности, вопреки очевидности отрицая явно противоречащие ему случаи, как, например, результаты скрещивания человеческих рас белой и черной, дающие, как известно, средний тип мулатов, квартеронов и т.д., а не представляющих ни явления доминантности, ни возвращения к чистым типам предков. Став на эту узкую точку зрения, мендельянцы свели общий важный закон случайных комбинаций расчлененных признаков (hybridation disjointe Нодена, segregation Дарвина) на совершенно второстепенный случай антагонизма между доминантами и рецессивами, хотя позднее сами стали от этого отрекаться. К тому же и самый факт вытеснения одного признака другим вовсе еще не разъяснен. Весьма вероятно, что способность, например, желтого цвета вытеснять зеленый принадлежит не самому желтому пигменту, а третьему, постоянно его сопровождающему телу. Далее выяснилось, что доминантность и рецессивность не постоянные свойства, а могут взаимно превращаться, и на этот раз выяснилось, что причинная связь явлений объясняется действием посторонних тел — ферментов.
Такова первая и основная задача изучения наследственности, как всегда заключающаяся в возможно систематической классификации явлений, подлежащих изучению. За нею и даже, как это нередко бывало в истории наук, одновременно с нею выдвигается, назревает другая потребность — объединения, объяснения совокупности фактов одной общей теорией. Но ни одна из предложенных до сих пор так называемых теорий наследственности не удовлетворяет требованию, которое, прежде всего, можно предъявить им, не может служить общей рабочей гипотезой, то есть орудием для направления исследований к открытию новых фактов, новых обобщений. Теории эти возникали еще в XVIII веке, но особенно многочисленны они стали с половины XIX века, толчком и образцом для чего послужила теория пангенезиса, предложенная Дарвином, от которой он сам позднее благоразумно отказался. Все они представляют одну и ту же попытку материального изображения преемственной передачи свойств от одного поколения другому. Все они в основе — только вариации на тему: потомство «плоть от плоти, кров от крови» своих предков; только с успехами наблюдения подставляются все более глубокие черты строения «клеточка от клеточки», «плазма от плазмы», «ядро от ядра», «хромозома от хромозомы» и т.д. (см. оплодотворение). А когда оказывалось недостаточно этих реальных материальных носителей наследственности, изобретались легионы воображаемых (геммуль, идиоплазм, зародышевых плазм, ид, идантов, детерминантов, генератуль, эргатуль, эргатин, пангенн, просто ген, мнем и т.д.), вся задача которых сводилась к тому, чтобы наглядно изобразить основной факт непосредственной материальной связи между частями организма и преемственной связи при смене поколений. Самой загадочной при этом обыкновенно представлялась возможность совмещения в ничтожном объеме зачатка каждого организма всех особенностей строения, обнаруживаемых его вполне развитой формой со всеми проявлениями ее деятельности.
Задача эта представляется бесконечно сложной, если ее понимать в действительном фактическом смысле раскрытия бесконечной цепи причин и следствий, связывающей наблюдаемую в данный момент форму с ее зачатком или, что все равно, в обратном направлении — с зачатком будущего поколения. Логически же основная задача сводится к объяснению связи между исчезнувшей уже причиной и ее отдаленным последствием, подход, ключ к разъяснению чего должно искать, вероятно, в несравненно более простых случаях так называемых последствий (Nachwirkungen), примеры которых доставляет физиология, особенно физиология растений. А если до времени довольствоваться словесными параллелями, аналогиями, метафорами, то эти сравнения должны удовлетворять основному логическому условию всякого объяснения: должны восходить от сложного к простому. С этой точки зрения должны быть, безусловно, отвергнуты такие словесные объяснения явлений наследственности, как пользующиеся в настоящее время значительным сочувствием попытки Герринга и в недавнее время Земона. Оба эти ученые на расстоянии каких-нибудь 40 лет выступили с теорией наследственности, основанной на сравнении ее с памятью. Но и фактическое, и словесное разрешение двух указанных проблем наследственности — задача физиологии, разрешаемая или прямыми опытами, или аналогией с искусственными явлениями, а, к сожалению, до сих пор задача эта оставалась почти исключительно достоянием морфологов, которые считают ее разрешенной, если им удалось морфологически связать одну форму с другой, видимую глазом с видимой под микроскопом, или невидимой, добавляя остальное тем легионом слов, которые заставляют только порой сожалеть, почему эти современные ученые еще так свободно владеют греческим языком, что могут так легко и свободно засорять память людей эфемерными созданиями своей филологической фантазии. Физиология, конечно, пойдет строго систематическим путем, и на место внешних, эмпирически связуемых форм, окраски и других признаков проникнет в их внутреннюю физико-химическую, причинную связь, как это на днях было сделано химиками, пришедшими на выручку мендельянцев и разрешившими непосильный для них факт превращения их доминантных форм в рецессивные и обратно (исследования Армстронга и Кибля над разноцветными примулами). И прежде всего, должно проникнуться мыслью, что причины могут быть потенциальные, а не непременно морфологические и вообще иного свойства, чем вызываемые ими следствия. Это нагляднее всего выражено в том факте, что источником всех пигментов являются бесцветные тела — хромогены и вызывающие их превращения ферменты. Точно так же и причины определенных форм могут лежать в известных свойствах аморфного вещества. Аналогию тому можно найти в опытах образования из двух жидкостей искусственных клеточек (клеточки Траубе), представляющих явления усвоения, роста и некоторые другие особенности живых клеточек, или в следующем простом опыте — фокусе — так называемом фараоновом змее: щепотка порошка, будучи подожжена, развивается в значительных размеров спирально извивающееся тело с определенным внутренним пузырчатым как бы клеточным строением. Причина этой вполне определенной и неизменно осуществляющейся формы лежит в свойстве порошка — гореть, образовать легкоплавкую, вследствие выделяющихся газов, пузыристую, быстро затвердевающую массу, — в результате дающую подобие превращения небольшого объема вещества семени в превышающий его в сотни раз объем ростка с его рыхлым клеточным строением. На основную загадку наследственности — кажущееся действие на расстоянии причины, уже не существующей, но существовавшей в прошлом, несомненно, прольет свет, как сказано, изучение явлений так называемого «последействия» вроде следующего. Финолог может показать два совершенно сходных вертикальных ростка, находящихся в абсолютно сходных условиях, и сказать, что один из них через несколько времени искривится вправо, а другой влево, и предсказание его в точности исполняется. Дело в том, что он сообщил им различную наследственность, различное прошлое, различно повлиял на ближайший период их истории — и, по существу безразлично, что период этот измерялся часами, а не годами или тысячелетиями, как в обычных явлениях наследственности; основное содержание явления — действие отсутствующей, но существовавшей в прошлом причины — одно и то же. Изучая все более и более сложные последействия, физиолог со временем доберется и до явлений наследственности. Точно так же и загадочность сосредоточения в ничтожном комке зародышевого вещества всех свойств будущего организма мы должны объяснять себе не существованием его в редуцированном морфологическом же состоянии (подобно известной теории emboitement, по образцу которой построены в основе и все подобные позднейшие теории), а в состоянии физико-химическом, потенциальном. А пока не удастся фактически проследить и хотя бы частично воспроизвести связь известных процессов через все промежуточные стадии развития, приходится довольствоваться логическими аналогиями, которые облегчали бы мысленное представление о природе этой связи. Такое представление может нам дать хотя бы ряд фотографических камер, поставленных одна за другой и воспроизводящих на своем матовом стекле то же изображение. Здесь мы усматриваем главное подобие явлений наследственности — периодическое воспроизведение сходных образов, чередующихся с местами, в которых невидимо, потенциально присутствуют эти образы, но в которых мы не могли бы их увидеть, — они должны развиться, и тогда только мы узнаем их вновь со всеми тонкостями их очертания и окраски. Такое сравнение, конечно, нас не далеко подвигает в фактическом познании, но исполняет свое назначение, подготовляя, приучая ум на более простых примерах к охвату более сложных задач.
Таблица V.
Помесь шести видов ивы попарно и трех помесей, вновь попарно, в помеси второго порядка.
(По Вихура).
Естественное самопечатание (Naturselbstdruck).
Когда Сеченов хотел сделать более доступным понятие физиологического механизма памяти, он его сравнил с только что открытым фонографом; никогда, конечно, физику не пришло бы в голову обратно объяснять простое устройство фонографа сравнением его с памятью.
В конечном выводе, мы должны ожидать разрешения задач наследственности от экспериментальной морфологии, в опытах наследования приобретенных свойств, но опытах, распространенных на более долгие периоды времени, помня, что и сама наследственность, в конце концов, может быть только «приобретенное свойство». А до той поры можно, пожалуй, освещать свой путь временными словесными аналогиями, но под непременным условием, чтобы они восходили к чему-нибудь реальному и простейшему, а не к фиктивному и более сложному, в роде модной теории «мнемонических протомер» с их «сублиминальными» и «супралиминальными» «энграммами» (Земон, 1904), памятуя известное изречение грозного бича старых схоластиков (Уильяма Оккамского): Entia non sunt multiplicanda praeter necessitatem (не следует без надобности размножать всякие сущности).
Литература: Darwin, «The Variation of Animals and Plants», 1868, втор. изд. 1875 (главы XI-XIX заключают всесторонний критический обзор основ учения о наследственности; русский перевод издания Лепковского, 1908); Darwin, «Cross and selffertilisation of Plants», 1876 (о пользе и роли в природе перекрестного оплодотворения); Naudin, «Sur l’Hybridité dans les végétaux», 1865 (Nouv. Arch, du Museum); Mendel, «Versuche über Pflanzenhybride», 1901 (первонач. 1865); Wichura, «Die Bastardbefruchtung im Pflanzenreich», 1865; Millardet, «L’hybridité sans croisement» (1894); Kerner, «Pflanzenleben» (в. 2, 1891); Costantin, «L’héredité acquise»; Henslow, «Heredity of acquiered characters in plants» (1908); Bateson, «Mendel’s principles of heredity» (1909); Gorrens, «Die Neuen Vererbungsgesetze», 1812 (есть русский перевод); А. Thomson, «Heredity» (1908); Е. Baur, «Einführung in die experimentelle Vererbungslehre», 1911 (есть русский перевод), Goldschmidt, «Einführung in die Vererbungswissenschaft», 1911 (русский перевод); Plate, «Vererbungslehre», 1912.
К. Тимирязев.
| Номер тома | 29 |
| Номер (-а) страницы | 611 |