Растение
Растение. 1. Ходячее понятие о растении резко противопоставляет его животному организму. Однако, современное состояние знания не оставляет никакого сомнения, что деление органического мира на два царства природы является условным, и что растительный и животный мир представляют собой лишь две мощных эволюционных ветви, имеющие общие исходные корни. Нет таких признаков, которые были бы свойственны только растению или только животному. Вместе с тем строительный материал, из которого слагается тело тех и других, является по существу тождественным (см. XX, 215/19).
Питание растений. Из всех особенностей, отличающих растительный организм от животного, за наиболее характерную и существенную нужно считать способ питания. Растения в отличие от животных обладают способностью вырабатывать, синтезировать сложные органические соединения из простых неорганических (автотрофный способ питания), причем особенно характерной чертой этого процесса является ассимиляция углерода (см. фотосинтез). Так как неорганические вещества поступают извне частью в газообразной форме, частью в растворах, то растения всасывают их своей поверхностью осмотическим путем. Животные организмы питаются только готовыми сложными органическими соединениями (гетеротрофный тип питания) и для переработки и усвоения их вводят таковые вовнутрь своего тела. Можно сказать, что и все строение растения и животного является в конечном итоге прямым или косвенным результатом приспособления «лаборатории организма» к способу и условиям питания.
Тем не менее и в этом отношении не находится действительных оснований для принципиального противопоставления растения и животного. Если способность растения самостоятельно синтезировать сложные вещества и является для него характерной, то в то же время переход к гетеротрофному питанию в виде сапрофитизма и паразитизма в растительном царстве (см. XXXI, 184/88) наблюдается в самых различных синтетических группах, от низших до высших. Особенно замечательны в том же отношении насекомоядные растения (см. XXIX, 648/51 и прил.), у которых гетеротрофный тип питания сопровождается появлением целого ряда черт животной организации. Сюда относятся: активное захватывание органической пищи, повышенная чувствительность и способность к движениям, образование пищеварительных полостей (рис. 1), повышенное выделение ферментов, тождественных с животными.
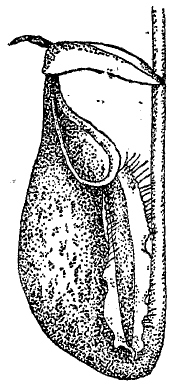
Рис. 1. Кувшинчик насекомоядного растения непентеса.
Заслуживает быть отмеченным, что современные насекомоядные растения представляют разорванные остатки явно вымершей биологической группы с большим числом монотипных (то есть имеющих всего один вид) родов (Dionaea, Ald ovandia, Cephalotus, Drosophyllum и др.). Это определенно указывает, что насекомоядные растения когда-то имели гораздо большее распространение, чем в настоящее время, и представляли как бы эволюционную попытку разрешения в крупном масштабе проблемы животного питания при помощи организационных возможностей, имеющихся у высших растительных организмов. Можно думать, что названная попытка находила какие-то благоприятные условия в соответствующую геологическую эпоху, позднее исчезнувшие. Кроме того, особенности некоторых современных австралийских росянок (см. XXIX, 651/52, приложение 8'/11') допускают мысль о возможности существования в прошлом насекомоядных растений с решительным преобладанием гетеротрофного питания над автотрофным, тогда как у большинства современных насекомоядных растений первое только дополняет второе и не является обязательным. С другой стороны, если в животном царстве вторичный способ питания, говоря вообще, выдерживается строго, то все же существуют простейшие организмы (например, Peridineae, многие Flagellata), которые зоологами обычно рассматриваются как животные, но которые обладают подобно растениям аппаратом, необходимым для самостоятельного синтеза. Одноклеточные организмы этого типа, по предложению Гэккеля, получили название протистов, и не остается никакой тени сомнения в том, что животный тип питания вырос, так сказать, на почве растительного.
Общие различия во внешней форме растений и животных, равно как отсутствие у первых подвижности, стоят в ясном соотношении с процессом ассимиляции углерода, как основной чертой растительного типа питания. Для растительных организмов характерно стремление к увеличению поверхности сравнительно с массой тела, выражающееся в ветвлении и образовании пластинчатых органов; для животных, наоборот, характерна концентрация массы и сведение поверхности тела до минимума, чем уменьшается сопротивление при движении. Однако, с одной стороны, у растений в связи, главным образом, с необходимостью ограничивать испарение (например, в пустынях) могут развиваться чисто геометрические формы тела — шары, цилиндры (у кактусов; см.), с другой — животные, обитающие в воде, могут сближаться с растениями по внешнему виду до неузнаваемости (зоофиты старых натуралистов, рис. 2). Это объясняется тем, что в воде, особенно в морях, плавает или взвешено много пищевого материала, падающего пассивно на дно или двигающегося мимо. Отсюда является возможность питания на счет его, не сходя с места, но увеличивая залавливающую поверхность подобно тому, как происходит улавливание частиц углекислого газа растений. Сближение внешнего механизма добычи пищи привело в соответствующих случаях к выработке и сходной внешней формы.
Рис. 2. Растениеобразное животное-гидрополип сертулярия.
Движения у растений. При животном типе питания подвижность, говоря вообще, составляет условие, необходимое для добычи пищи. При растительном способе необходимость в активных движениях уже сама по себе отпадает но, кроме того, в самом процессе ассимиляции кроются условия, повлекшие за собой в ходе эволюции растительного царства ограничение подвижности до минимума. Процесс ассимиляции углерода сопровождается образованием веществ (растворимых углеводов и др.), обладающих сильным осмотическим притяжением веды. На этой почве внутри растительной клетки происходит накопление последней с образованием полостей (вакуоль), наполненных клеточным соком, и вместе с тем возникает сильное внутриклеточное давление (тургор), стремящееся разорвать протоплазму клетки. Поэтому специализация растительной клетки как синтезирующего аппарата в ходе эволюции сопровождалась выработкой приспособления для противодействия тургору - защитной клеточной оболочки, которая со своей стороны сделала свободную подвижность невозможной. Опять-таки среди протистов можно видеть как бы отдельные ступени, которыми шло все более и более полное замыкание живого содержимого в твердую оболочку и утрата подвижности. У перидиней (см. Х, 555/56, и табл. I, 1) оболочка состоит из отдельных пластинок, примыкающих друг к другу краями, и совмещается с жгутиками, позволяющими еще полную подвижность. У диатомей (см. Х, 555/56 и табл. I, 2-5, и XVIII, 454) она состоит из двух половинок, которые наподобие створок заходят краями друг за друга. У некоторых жгутиковых (см.) оболочка облекает тело клетки почти кругом, но все же имеет отверстие, через которое выходят жгутики. Только путем превращения оболочки в совершенно замкнутый прочный мешочек была достигнута возможность максимальной продуктивности синтеза при наличности внутриклеточного давления в несколько атмосфер. Вместе с тем, очевидно, ход эволюции в этом направлении шел решительно в сторону отказа от подвижности.
Тем не менее, способность реагировать на внешние раздражения то медленными, мало заметными, то более быстрыми изгибами и перемещениями частей не только свойственна каждому растению, но и лежит в основе всей его механики жизни. Она определяет рост в строго постоянном, явно приспособленном к условиям физиологической деятельности, направлении стебля и корня (положительный и отрицательный геотропизм), световую установку листьев (фототропизм), разыскивание влаги в почве корнями (гидротропизм), обвивание вокруг подпорки усиков и вьющихся растений (тигмотропизм) и проч. (см. тропизмы). Иногда же чувствительность и способность движения частей у растений приобретает настолько резко выраженные формы, что поведение растений ничем не отличается от поведения даже высших животных. Так, насекомоядное растение мухоловка (см. XXIX, 651/52, приложение 11'/12') с такой быстротой захлопывает половинки своего листа, что насекомое не успевает улететь и оказывается пойманным. Стыдливая мимоза (см. ХХVIII, 671/72) под влиянием сотрясения почти моментально складывает свои листья и совершенно изменяет свой внешний вид (рис. 3). С другой стороны, под влиянием анестезирующих веществ (хлороформа, серного эфира) она переходит в состояние усыпления и нечувствительности. Конечно, у растений явления чувствительности и двигательной реакции не идут дальше примитивных форм рефлекса, не сопровождаясь какими-либо внутренними психическими состояниями (см. чувствительность растений). Но и у животных сознание, без сомнения, является свойственным только высшим этапам эволюции и развивающимся, как приспособление, из сложного переплета простейших «условных» рефлексов.
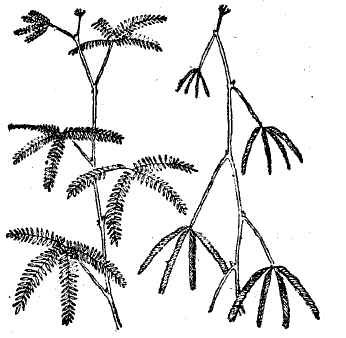
Рис. 3. Стыдливая мимоза в состоянии покоя (слева) и потревоженная (справа).
Единство жизни. Внутреннее единство растений и животных выясняется самым бесспорным образом во всем ходе новейшего естествознания на основании глубокого сходства как в общих принципах их строения, так и в физиологических процессах и характере действующих в них химических соединений. Одним из замечательнейших из числа многочисленных относящихся сюда примеров внутреннего тождества, бесспорно, является почти полное сходство процесса сложного деления клеточного ядра (см. кариокинез, XXIII, 492/94, и XX, 224/26), которое обнаруживается от низших до самых высших животных и растительных организмов. Не менее поразительно присутствие тождественных или очень близких химических веществ как в растительном, так и в животном организме, число которых в свете современной биохимии все растет и растет. Так, характернейшее вещество растительного царства — зеленый пигмент, хлорофилл, оказался, по исследованиям Ненцкого и Мархлевского, химически близко родственным с пигментом крови животных — гемоглобином. При этом даже и роль их в химизме газообмена, по новейшим исследованиям Вилльштеттера, оказывается очень близкой. Другое характернейшее вещество растений, клетчатка, вдруг появляется у оболочников (Tunicatа), хотя и отдаленных, но прямых родичей позвоночных животных. И обратно, хитин, вещество, распространенное в целых больших группах животных, имеет свою близкую химическую пару в оболочках грибов. Характерный для животных продукт обмена веществ — мочевина — найден опять-таки у грибов. И в растительном, и в животном организме работают одни и те же или почти тождественные ферменты. Белковый обмен у животных и у растений сопровождается глубоким расщеплением белковой частицы до амидокислот с обратным синтезом из последних частиц белка.
2. Растительная клетка. Типическая растительная клетка (см.) отличается от животной рядом признаков, — прежде всего присутствием внутри протоплазмы особых белковых телец-пластид (см. хроматофоры).
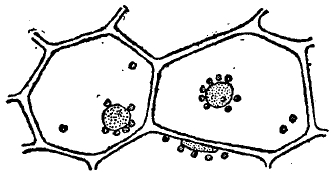
Рис. 4. Лейкопласты вокруг клеточного ядра.
Они наиболее резко выступают (рис. 4), когда они окрашены, чаще всего хлорофиллом (см.), в зеленый цвет (хлоропласты, хлорофилльные зерна), реже в другие цвета — желтый, оранжевый (хромопласты), но пластиды могут быть и совершенно бесцветными (лейкопласты) и тогда плохо заметными и легко ускользающими от наблюдения. Относительно способа возникновения пластид, согласно с взглядами Шимпера, считалось, что они, как и ядра, образуются только делением (путем простой перетяжки) раньше существовавших пластид. В новейшее время ряд исследователей высказывается в пользу возникновения пластид из так называемых хондриосом (см.). Хлоропласты и лейкопласты высших растений имеют вид округлых или эллиптических телец, находящихся в значительном количестве внутри протоплазмы. По своей физиологической роли пластиды являются химическими очагами, имеющими теснейшее отношение к синтезу углеводов. В зеленых пластидах происходит отложение первичного крахмала, возникающего в процессе ассимиляции; в лейкопластах крахмал отлагается на вторичных местах, в качестве запасного — в клубнях, семенах и проч. Другой особенностью растительной клетки является уже упоминавшееся ранее возникновение в ней полостей (вакуоль), наполненных клеточным соком. Состав клеточного сока весьма разнообразен и сложен (см. растительные соки). Из важнейших веществ, которые в него входят, надо назвать, прежде всего, растворимые минеральные соли. Они проникают извне осмотическим путем, и содержание его в известных пределах отражает химический состав окружающей среды. Так, на солончаковых почвах и клеточный сок богат соответствующими солями.
Но поступление солей в клетку в сильнейшей степени регулируется избирательной способностью клетки. Благодаря ей, в растениях могут накапливаться в большом количестве вещества, содержание которых в окружающем растворе может быть совершенно ничтожно. Так, одноклеточные диатомеи имеют оболочку, пропитанную кремнеземом, хотя этот последний растворим в воде лишь в виде ничтожных следов. Клетка как бы выбирает из растворов, что ей нужно. Избирательная способность легко объясняется тем, что вещества, которые принимают близкое участие в химизме клетки, по поступлению в нее сейчас же подвергаются тому или иному химическому видоизменению. Вследствие этого постоянно поддерживается разность концентрации внутри клетки и во внешней среде. Наоборот, вещества, которые чужды химизму клетки, могут поступать в нее самое большее до выравнивания концентраций. Вопрос об избирательной способности клетки теснейшим образом связан с вопросом о физиологической роли отдельных элементов. Однако, в этой области далеко не все ясно. Некоторые элементы, как железо, встречаются в растениях в минимальном количестве и все же безусловно необходимы. Наоборот, например, кремний в некоторых растениях содержится в большом количестве, а между тем соответствующие растения (например, злаки) удается искусственно выращивать вполне успешно на питательных средах, совершенно лишенных названного элемента. Число элементов, которые обнаруживает анализ золы растений в различных случаях, весьма велико. Но методом искусственных культур (см. водные культуры) найдено, что, безусловно, необходимыми для растений являются только (кроме углерода, кислорода и водорода) азот, фосфор, сера, калий, кальций, магний и железо. Роль их в химизме растений пока не может считаться достаточно выясненной. Азот (см. ассимиляция азота), фосфор, сера входят непосредственно в состав белковой частицы. Железо и магний имеют ближайшее отношение к образованию хлорофилла и его деятельности (см. хлорофилл). Калий имеет ближайшее отношение к жизнедеятельности протоплазмы, хотя точнее о его роли можно говорить только гадательно. Кальцию приписывается важное значение в процессах расщепления и синтеза белковой частицы (нейтрализация щавелевой кислоты). Кроме неорганических соединений, в состав клеточного сока входят органические кислоты (щавелевая, лимонная, яблочная и др.), дубильные вещества, пигменты. Из пигментов особенное распространение имеет антокиан (см.), или антоциан, придающий соответствующим частям растений окраску от розовой и фиолетово-красной до синей. Оттенок окраски антоциана зависит от химической реакции клеточного сока — кислой или щелочной. Под влиянием антоциана листья растений нередко бывают красного цвета. Однако, стоит прокипятить такой лист, и он зеленеет: антоциан легко диффундирует и исчезает из убитых клеток, а хлорофилл остается.
Во вполне выросшей растительной клетке обыкновенно или образуется одна сплошная вакуоля, растягивающая протоплазму до тонкого малозаметного слоя, прижатого к оболочке, или же по существу такая же вакуоля пересекается более или менее многочисленными тяжами протоплазмы. Что клеточный сок растягивает с силой протоплазму, легко обнаруживается при прибавлении к микроскопическому препарату какой-либо жидкости (например, глицерина) или раствора (сахара, селитры), сильно отнимающего воду. Тогда наступает явление плазмолиза: протоплазма отстает от оболочки и съеживается в шаровидное тело. Также явление плазмолиза клеток лежит в основе увядания (клетки теряют воду испарением при недостаточном снабжении ею). Это внутриклеточное давление в растительной клетке, вызывающее наряженное состояние целлюлозных оболочек (тургор), играет важную роль в росте клеточки. Благодаря ему растительная клетка вообще крупнее животной, а иногда (сифоновые водоросли, см. Х, 557, 548, табл. I и II, 24-30) может достигать исключительных размеров в несколько сантиметров. Внутриклеточное давление часто достигает 3-5 атмосфер, а в отдельных случаях 10 и более атмосфер (у растений, живущих в условиях крайней сухости). Тургор тканей вместе с тем придает устойчивость сочным молодым органам.
Наконец, третьей характернейшей особенностью растительной клетки является уже упоминавшееся присутствие твердой оболочки. Эта последняя в молодых клетках обыкновенно состоит из клетчатки (см. XXIV, 353/54), или целлюлозы, углевода, близко родственного с крахмалом (с формулой С6Н10О5). Для знакомства с чистой клетчаткой может служить очищенная вата, которая представляет собой оболочки волосков, покрывающих семена хлопчатника. В окончательном состоянии оболочка может подвергаться различным химическим видоизменениям. Особенно большое значение имеют процесс одеревенения и процесс опробковения. Одеревеневшие оболочки отличаются большей прочностью и меньшей растяжимостью, чем целлюлозные. Они дают иные микрохимические реакции (с иодом и серной кислотой, флороглюцином и соляной кислотой и др.; см. древеснение, XIX, 75/76). Процессу одеревенения подвергаются у высших растений определенные группы клеток, главным образом проводящие воду части сосудисто-волокнистых пучков (см. древесина) и механические ткани. Клетки с одеревеневшими оболочками составляют основную массу вещества стволов деревьев и определяют их техническое применение. Растительные продукты с большим содержанием одеревеневших оболочек малопригодны для питания животных. При опробковении оболочки как бы пропитываются жирообразным веществом и делаются непроницаемыми для жидкостей и газов. Поэтому клетки, у которых опробковение охватывает оболочку на всей поверхности, совершенно теряют свою жизнедеятельность. Опробковению подвергаются почти исключительно клетки, лежащие на самой поверхности органов в связи с их ролью защиты растений от испарения (см. транспирация) и других неблагоприятных условий. Степень развития может колебаться от тончайшей микроскопической пленки (кутикулы), имеющейся в кожице каждого сухопутного растения, до мощных отложений пробковой коры, развивающихся у древесных пород (см. феллоген). Толщина клеточной оболочки бывает в разных случаях весьма неодинакова. У механических клеток, то есть предназначенных служить для укрепления тех или других органов растений, утолщение оболочки может заходить так далеко, что полость клетки почти сходит на нет (см. стереиды). На утолщенных оболочках обыкновенно хорошо видна слоистость, параллельная поверхности. В оболочках имеются всегда тонкие места — продушины, или канальцы, приходящиеся в смежных клетках как раз друг против друга и служащие для тесного контакта между протопластами клеток. Тончайший слой оболочки, который находится на границе соприкосновения двух смежных растительных клеток (пограничная пластинка), состоит из особого пектинового вещества, близкого к углеводам, но менее стойкого, чем остальная оболочка. Поэтому в этом слое может происходить разъединение клеток, как естественное (например, в мякоти зрелых плодов), так и искусственное (мацерация; например, при кипячении с едким кали или с азотной кислотой и бертолетовой солью).
Среди разнообразных представителей типа протистов можно наблюдать отдельные этапы, по которым шла специализация растительной клетки, не только по отношению к выработке твердой оболочки, но и со стороны вообще клеточного аппарата. В этом отношении особенно замечательны сине-зеленые водоросли (см.) — Cyanophyceae, соединяемые обычно на основании общего признака — отсутствия обособленного клеточного ядра — вместе с бактериями в одну систематическую группу дробянок (см.). По присутствию пигмента, близкого к хлорофиллу, и твердой оболочки они являются вполне выраженными растительными клетками, но у них отсутствуют еще пластиды, и ассимиляторный пигмент диффузно распространен в протоплазме. Вместе с тем у них нет и центральной вакуоли. Еще более раннюю ступень представляют бактерии, так как у них химизм питания представляет чрезвычайное разнообразие. Среди бактерий встречаются как бы попытки синтеза органического вещества путем использования иных форм энергии, чем световая (хемосинтез, см.), причем отсутствует и ассимиляторный пигмент.
Общими обязательными частями растительной и животной клетки являются протоплазма и ядро (см. XX, 211 сл., 221 сл.). Первая представляет полужидкое прозрачное вещество, в котором даже при сильнейших увеличениях удается видеть только мельчайшие зернышки — микрозомы. Отношение их к протоплазме, вероятно, не всегда одинаковое. Только на поверхностях соприкосновения с окружающей средой и с клеточным соком протоплазма отличается большой плотностью и совершенной однородностью, без зернышек (так называемый кожистый, или стекловидный слой). В ряде случаев (в клетках листьев водяных растений, валлиснерии и элодеи, в жгучих волосках крапивы и некоторых других) наблюдается движение протоплазмы внутри оболочки, или вращательное (вдоль стенки), или струйчатое (вдоль тяжей протоплазматической сетки). Клеточное ядро в растительных клетках обыкновенно имеет вид округлого или несколько вытянутого бесцветного тельца с одним или несколькими блестящими зернышками — ядрышками. При помощи метода окрашивания обнаруживается строение ядра из слабо окрашивающего остова — линина, и сильно окрашивающегося вещества — хроматина.
3. Слоевцовые растения. Древнейший в эволюционном смысле тип растительных организмов представляют водоросли (см.). Под этим названием объединяются многочисленные растительные организмы, обитающие, за немногими исключениями, в воде и характеризующиеся слабой как внешней, так и внутренней дифференцировкой тела или даже полным отсутствием дифференцировки (слоевцовые растения). В пределах их можно наблюдать все переходы от свободно живущих микроскопических организмов, состоящих всего из одной клетки, до крупных многоклеточных. Тело водорослей лишено характерного для высших растений деления на резко морфологически отграниченные органы: листья, стебель, корни. Оно имеет вид ветвящихся пластинок или кycтиков, причем у водорослей распространен дихотомический, или вильчатый способ ветвления (верхушка ветви каждый раз делится на две новых расходящихся ветви), несвойственный высшим растениям (см. XII, 161/62). Пресноводные водоросли имеют обычно вид тончайших зеленых нитей (нитчатки). В анатомическом строении водорослей обнаруживаются лишь слабые намеки на образование обособленных тканей: у них почти полностью отсутствует сосудистая система, слабо выражена покровная ткань и проч.
Некоторые простейшие водоросли (диатомеи, сине-зеленые) прямо еще несут черты протистов, но и все вообще водоросли обнаруживают в строении клеток целый ряд особенностей, свидетельствующих о близости их к корням эволюции. Так, строение ядра у водорослей в разной степени уклоняется от обычного. При кариокинезе у них наблюдаются свойственные животным клеткам, но отсутствующие у высших растений центрозомы (см. XXIII, 494). Нередки многоядерные клетки. Тип пластид представляет замечательное разнообразие (см. Х, 550). Наконец, и ассимиляторный пигмент у водорослей представляет гораздо большее разнообразие, чем у высших растений. У них фигурирует не только зеленый хлорофилл, но даже чаще другие пигменты — бурые, красные, представляющие, вероятно, сложные комплексные соединения хлорофилла. Оболочки клеток водорослей обнаруживают широкое распространение у них процесса ослизнения.
Клетки водорослей сами по себе обыкновенно неподвижны, но способность к движению их живого содержимого сохраняется у большинства в весьма своеобразной форме, опять-таки связующей их с протистами. Протопласт водорослей, при известных условиях целиком (например, у вошерии, Vaucheria) или предварительно разбившись на большее или меньшее число отдельных телец (наиболее обычный способ), покидает оболочку материнской клетки и начинает двигаться в воде. Такого рода временная подвижная стадия водорослей называется зооспорой (см. Х, 551/52). Половое размножение у водорослей совершается также существенно иначе, чем у выше организованных растений и опять-таки представляет целый ряд значительно отличающихся друг от друга вариантов (например, копуляция у спирогиры, см. Х, 552). Вместе с тем у водорослей можно проследить все ступени эволюции полового процесса: он начинается с изогамии, то есть слияния совершенно сходных между собой гамет, и кончается типическим половым размножением — оогамией, когда сливающиеся клетки специализируются на определенном физиологическом поведении и принимают совершенно различную внешнюю форму. Одна из них – яйцевая клетка, отличается сравнительно крупными размерами и обильной густой протоплазмой, но лишена подвижности и имеет обычно круглую форму. Другая – мужская, живчик, сперматозоид, наоборот, очень мелка, состоит преимущественно из ядерного вещества и бедна питательными веществами, но обладает жгутиками и ресничками и легко подвижна. Она является оплодотворяющей клеткой (см. Х, 553/54).
Половой процесс у водорослей мы захватываем как бы in statu nascendi. Вместе с тем у водорослей удается экспериментально очень легко вызывать замену полового размножения бесполым и обратно. При изменениях состава окружающей среды гаметы могут вести себя как вегетативные зооспоры и развиваться дальше без предшествующего слияния. Как показали опыты Клебса, ухудшение условий питания дает толчок к половому размножению, хорошие условия питания благоприятствуют вегетативному размножению. В руках экспериментатора находится полная возможность управлять ходом развития организма и сменой его последовательных состояний, которая на первый взгляд кажется чем-то строго зафиксированным и всецело определяющимся внутренними причинами.
У водорослей нередко наблюдается ясно выраженное чередование полового и бесполого поколения: оплодотворенная яйцевая клетка сначала дает клеточное тело, размножающееся вегетативно (зооспорами или иначе), и только эта новая генерация повторяет половое размножение. Благодаря чередованию поколений, один акт оплодотворения дает сразу начало большому количеству особей. Этим вносится корректив к некоторой необеспеченности полового процесса. Особенно выраженные формы чередование поколений принимает у некоторых бурых водорослей (Laminariaсеае), сближаясь с картиной того же явления у высших споровых. Половое поколение имеет вид небольшого ветвистого нитевидного тела (ср. также Х, 554/55). Вместе с тем уже у водорослей намечается связь чередования поколений с редукционным делением, то есть тем видоизменением кариокинетического процесса, которое ведет к уменьшению числа хромосом вдвое. Этим устраняется прогрессивное увеличение числа хромосом, к которому должно было бы вести слияние мужского и женского ядра при половом размножении ср. XX, 232/33). В вышеупомянутых случаях чередования поколений бесполая генерация имеет полное число хромосом (принято обозначать 2 х). При образовании зооспор происходит редукционное деление, и развивающаяся из зооспоры половая генерация имеет клетки с вдвое меньшим числом хромосом (х). После слияния половых клеток снова восстанавливается 2 х хромосом.
Значительные уклонения в процессе оплодотворения среди водорослей имеют место у багрянок (Rhodophyceae) и у харовых (Characeae). У первых из них мужские клетки (равно как и клетки, соответствующие зооспорам других водорослей, называемые здесь тетраспорами) неподвижны и пассивно приносятся водой к женскому аппарату — карпогонию (см. Х, 559/60, табл. II, 41). Этот последний представляет оогоний, снабженный волосовидным придатком (трихогиной), приспособлением, направленным к увеличению улавливающей поверхности оогония (аналогично рыльцу цветковых) для большего обеспечения попадания на него мужских клеток (спермациев). Спермации прилипают к трихогине, между содержимым спермация и трихогиной устанавливается прямое сообщение, и ядро спермация по трихогине проникает в основную часть карпогония, где находится яйцевая клетка. В результате оплодотворения у багрянок происходит сначала образование особых ветвистых нитей, представляющих собой опять-таки бесполую генерацию, и только эта последняя приносит карпоспоры. Вместе с тем она является — 2 х-генерацией, как бы паразитирующей на половой х-генерации.
У харовых (см. Х, 559/60, табл. II, 31 А, В, С) оогоний окружен снаружи более прочным вместилищем, образующимся вследствие разрастания и тесного смыкания кольца клеток из основания оогония. Их сперматозоиды напоминают сперматозоиды мхов, но по способу образования сильно отличаются от всех других смежных растений. Современные харовые и в других отношениях стоят обособленно от остальных водорослей, представляя собой последние остатки какой-то, пока загадочной, вымершей группы. Они напоминают собой более высокоорганизованные растения. Вообще следует признать, что обширный мир простейших растительных организмов, объединяемых термином «водоросли», представляет собой в действительности несколько совершенно самостоятельных линий эволюционного развития.
Грибы. К водорослям теснейшим образам примыкают две группы растительных организмов: грибы (см., Fungi) и лишайники (см., Lichenes). Первые из них можно рассматривать как непосредственную ветвь водорослей, перешедших к гетеротрофному питанию — сапрофитному и паразитному, и претерпевших процесс глубокого приспособления к способу питания. Соответственно у них клетки утрачивают ассимиляторный пигмент и пластиды. Образование центральной вакуоли слабо выражено, вследствие чего клетки грибницы, или мицелия, отличаются незначительной толщиной и имеют вид вытянутых тонких нитей (см. XVII, 95/96, и табл.). Замечательным образом и оболочка у грибов обычно состоит из хитина (см.), вещества, свойственного животному царству. Особенно ясна родственная связь с водорослями у низших грибов — фикомицетов (phycomycetes), или грибов-водорослей (см. XVII, 99/103). Многие из них живут в воде или нуждаются, по крайней мере, в избытке влаги. Вместе с тем у них наблюдается образование подвижных зооспор и ясно выраженный половой процесс по типу водорослей. У одних фикомицетов (у Oomycetes) имеет место образование оогония и антеридия, у других (у Zygomycetes) — конъюгация, или слияние содержимого двух сходящихся концами веточек мицелия. Как раз один из представителей оомицетов — грибок, развивающийся на трупах мух, попавших в воду, сапролегния (Saprolegnia mixta), послужил вместе с водорослями материалом в замечательных опытах Клебса для экспериментального изучения вопроса о зависимости способа размножения от влияния внешних условий (см. XXIX, 620/22). Так, оказалось, что у сапролегнии по произволу можно иметь: 1) непрерывный деятельный рост, культивируя ее во всех хороших питательных растворах до тех пор, пока имеется налицо свежее неизмененное питание; 2) полное превращение мицелия в зооспорангии, если перенести хорошо питающийся мицелий в чистую воду: 3) рост одновременно с образованием зооспор, при культуре мицелия на агар-альбумине, находящемся в текучей воде; 4) сначала рост, затем оживленное образование половых клеток, при культуре в растворах лейцина или гемоглобина; 5) сначала рост, затем образование зооспор и, наконец, образование половых клеток или при культуре на мертвых насекомых, или если культивировать мицелий на желатине - мясном экстракте и затем перенести его в воду; 6) рост и одновременное образование зооспорангиев и половых клеток, при культуре в воде с прибавлением фибрина. Эти опыты позволяют объяснить поведение сапролегнии в ее обычных условиях появления на трупах мух. Ее мицелий сначала развивается в самом трупе, в условиях избыточного питания, и дает картину оживленного роста. Органические вещества из трупа диффундируют в окружающую воду, и вокруг трупа образуются концентрические жидкости с постепенно убывающей концентрацией питательных веществ. Мицелий начинает расти в окружающую жидкость и достигает почти чистой воды. Тогда начинается образование зооспорангиев. Определенные внешние условия вызывают и строго определенную реакцию организма.
Тот факт, что фикомицеты отличаются от высших грибов одноклеточной грибницей, дает право непосредственно сближать их с сифонниковыми водорослями, тело которых также внутри лишено перегородки.
Высшие грибы (см. XVII, 103/09, 97/99) — сумчатые (Ascomycetes) и базидиальные (Basidiomycеtеs), — являются уже всецело наземными организмами, но скрывающимися обычно в вегетативном состоянии в питательном субстрате и поэтому не обладающими специальными приспособлениями для защиты от высыхания, оставаясь на низком уровне организации. Вместе с тем, с одной стороны, у относящихся сюда грибов образование зооспор заменяется образованием лишенных подвижности и более выносливых спор, которые представляют большое разнообразие по способу своего развития нередко даже у одного и того же вида (наиболее яркий пример — ржавчинники). С другой стороны, половой процесс у высших грибов до крайности редуцируется (см. XVII, 101). Сравнительно легче он поддается обнаружению у сумчатых грибов, где он сводится к слиянию ядер двух тесно сближенных веточек гиф. У базидиальных грибов редукция идет еще дальше, и в конце концов от полового процесса остается только след в виде слияния парных ядер, находящихся в одной и той же клетке. Тем не менее, образование типических спор у высших грибов — аскоспор и базидиоспор — связано с предшествующим половым процессом, и таким образом у высших грибов также имеет место смена половой и бесполой генерации.
Лишайники (см.) представляют собой единственную в своем роде группу организмов, построенных на началах симбиоза (см.) и состоящих из гриба (обычно сумчатого) и водоросли (зеленой или сине-зеленой). Грибница образует бесцветную основу гриба, в петлях которой находятся клетки водоросли. Совместное существование их дает известные выгоды тому и другому организму. Гриб использует водоросль как источник питания и как бы поедает ее, водоросль же под защитой грибницы получает возможность существовать в условиях, необычных для водорослей и отличающихся большой сухостью. Таким образом, лишайники представляют собой как бы эволюционную попытку простейших растительных организмов перейти к существованию на суше без тех организационных возможностей, которые достигнуты были путем медленного приспособления высшими растениями. Способ размножения лишайников определяется их грибным компонентом.
Общая тенденция растительных организмов к развитию возможно большей ассимиляционной поверхности разнообразно проявляется и среди водорослей, приводя иногда к большому внешнему сходству с высшими растениями (например, у саргассовых бурых водорослей). Она сказывается даже в пределах одиночной клетки. Так, морская зеленая водоросль из сифонниковых — каулерпа (см. Х, 559/60, табл. II, 29) представляет гигантскую одиночную клетку, величиной с ладонь и больше, распадающуюся на части, соответствующие листьям, стеблям и корням.
Тот факт, что водоросли как бы застыли на низкой ступени внешнего и внутреннего расчленения тела, объясняется их исконным обитанием в воде, где условия жизни во многих отношениях более легки и просты, чем на суше, и не требуют выработки особых структурных приспособлений. Так, при постоянном обитании в воде отпадает опасность высыхания, а вместе с тем необходимость защитных покровов и проводящих воду трубок. Благодаря большой плотности среды, в воде почти не требуется специальных приспособлений для механической поддержки частей организма в пространстве. Поступление газов и минеральных растворов при водном обитании может происходить через всю поверхность растений. Только в воде возможны размножения при помощи зооспор, равно как и простейшие формы полового процесса, когда половые клетки самостоятельно находят свою пару вне родительского организма.
4. Переход растительного царства от жизни в воде к жизни на суше потребовал глубокой реорганизации всего строения растения и мог произойти только постепенно. Перед растением встала, как основная задача, защита от высыхания и обеспечение водой и минеральными солями из почвы. Отсюда, естественно намечается характерное для наземных растений строение из двойной системы усвояющих органов — надземных листьев и подземных корней, и связующего их остова — стебля. Одновременно для разрешения той же задачи потребовалась выработка внутреннего расчленения тела на ткани. Из последних у растения наиболее характерно оформлены — покровная ткань, регулирующая потерю влажности и газообмен с окружающей средой, проводящая ткань, служащая для более быстрой подачи воды и передвижения питательных веществ в места потребления и отложения запасов, и механическая ткань, сообщающая необходимую прочность органам растений. Строение клетки приобретает значительное однообразие. Подвижность утрачивается до максимума. В половом размножении яйцевая клетка остается на материнском растении и глубоко скрыта внутри защищающего от высыхания вместилища, проникновение же к ней мужских клеток осуществляется путем сложных приспособлений (внутреннее оплодотворение).
Мхи. Типом растительных организмов, связующих водоросли с более характерно выраженными обитателями суши, являются мхи (Bryophyta, см. мховые). Они распадаются на два класса: печеночники (см.) — Hepaticae и листостебельные мхи — Musci (см. мхи). Первые из них в своих простейших представителях могут быть еще отнесены вполне к слоевцовым растениям и представляют собой зеленые, дихотомически ветвящиеся пластинки. Они лежат непосредственно на влажной земле (иногда на поверхности водоемов) и выпускают снизу лишь корнеобразные волоски (ризоиды; см. XXXII, 147/48, табл. I, 2 и 8). Листостебельные мхи по внешности являются уже миниатюрными копиями высших растений и состоят из облиственных стебельков, выпускающих при основании ризоиды (см. XXIX, 455/56, табл. I,1 и др.). Все части мхов, однако, имеют незначительные размеры и нежны. Их листья чешуевидны и под микроскопом оказываются состоящими всего из одного слоя клеток. Точно также стебли в толщину не превышают какого-нибудь миллиметра. Образование тканей в их строении лишь намечается в виде слабо обособленной кожицы на поверхности и тяжа вытянутых клеток в середине, являющихся зачаточной сосудистой системой. Соответственно с слабым развитием тканей и органов всасывания, мхи никогда не достигают крупных размеров и в своем обитании связаны с условиями, характеризующимися обильным увлажнением, обитая на болотистых и лесных почвах.
У мхов, в очевидной связи с менее благоприятными условиями влажности, уже совершенно отсутствует вегетативное размножение зооспорами. В половом размножении точно так же наблюдается ряд особенностей, стоящих в явном соотношении с меньшей обеспеченностью водой. Яйцевая клетка мхов остается до конца на материнском растении и находится в особом вместилище — архегонии (см.), имеющем вид бутыльчатого тела, стенки которого состоят из одного слоя клеток. Антеридии (см.) представляют собой многоклеточные булавовидные тела, дающие из каждой клетки (кроме поверхностного слоя) спирально изогнутый сперматозоид с двумя жгутиками. Для оплодотворения необходимо, чтобы между входным отверстием архегония и антеридием находился соединительный слой воды, по которому сперматозоиды и проникают активно в архегонии (см. XXIX, 456/57, и XXXII, 149/50). Исследования Пфеффера показали, что из архегония выделяется вещество (у листостебельных мхов, вероятно, сахар, у печеночников — белкового характера), которое, диффундируя в воду, действует на сперматозоиды как раздражитель (хемотаксис), направляющий их движение по направлению к устью архегония. Развитие мхов связано с резко выраженным чередованием поколений: в результате оплодотворения из яйцевой клетки развивается особое тело — спорогон, являющееся бесполым поколением, приносящим лишь споры. Спорогон обычно делится на длинную, тонкую, как щетинка, ножку и коробочку (спорангий), отверстие которой окружено красивой зубчатой окраиной (перистом) и снабжено крышечкой. Внутри коробочки образуются путем деления многочисленные споры (по 4 из каждой материнской клетки). Из споры вырастает уже снова половое поколение. Спорогон при поверхностном взгляде кажется непосредственным продолжением полового поколения мха, образующим с ним одно целое. Все клетки спорогона имеют 2 х хромосом. При образовании спор происходит редукционное деление, вследствие чего все клетки полового поколения мха, развивающегося из споры, имеют х хромосом. Замечательно, что при прорастании споры сначала развивается нитевидное образование (протонема), которое по внешности до неузнаваемости походит на нитевидную водоросль (биогенетический закон), отличаясь, однако, от этой последней косо стоящими перегородками.
5. Высшие споровые растения. Дальнейший этап эволюции растительного мира в сторону завоевания суши представляют высшие споровые, или папоротникообразные (см.), куда в современном растительном покрове относятся собственно папоротники, хвощи и плауны. По внешнему и внутреннему строению вегетативных органов они стоят уже очень близко к цветковым, отличаясь от них лишь во второстепенных подробностях.
Более мощное развитие всех органов, в особенности корней, обладание сосудистой системой, образование покровной ткани, снабжение листьев проводящими пучками и проч. позволяет им, в отличие ото мхов, менее зависеть в условиях обитания от избытка влаги и достигать размеров целых деревьев. Однако, анатомическое строение споровых значительно уступает по своей приспособительной гибкости цветковым растениям. У них преобладает тип лестничных водоносных сосудов, притом не сплошных трубок, а лишь сильно вытянутых клеток (трахеид). Слабо выражена способность к вторичному утолщению. Жилки листьев дают мало анастомозов, отчего снабжение листа водой легко нарушается. Чешуйчатые же листья хвощей и плаунов имеют всего одну среднюю жилку. В связи с меньшей приспособляемостью высшие споровые боятся крайних условий температуры и сухости и являются детищами влажного теплого климата. Еще более зависимость от влажности у них проявляется при половом размножении. Для осуществления оплодотворения они так же нуждаются в воде, как и мхи. Вместе с тем у них чередование поколений принимает еще более резко выраженную форму, чем у мхов: индивидуальное развитие особи у высших споровых распадается в полном смысле слова на два самостоятельных существования. В отличие ото мхов половое поколение высших споровых (х-генерация), называемое заростком (гаметофит), гораздо слабее, чем бесполое, (2х-генерация) и по своему внешнему виду, незначительным размерам и простоте строения напоминает слоевцовые растения. У папоротников заросток обыкновенно имеет форму сердцевидной зеленой пластинки (см. XXXI, 133), лежащей на земле и выпускающей снизу ризоиды. На нижней же стороне ее находятся антеридии и архегонии, сходные с соответствующими органами мхов, но более упрощенные и погруженные основанием в ткань заростка (см. XXXI, 131/32, табл. II, 8 и 16). У хвощей заросток имеет вид еще более мелких курчавых пластинок (см. XLV, ч. 2, 497/98, табл. хвощи, 9, 10), у плаунов заросток, лишенный хлорофилла, клубнеобразный, скрытый в почве (см. XXXII, 336, рис., А и В). Процесс оплодотворения у высших споровых происходит совершенно сходно со мхами: сперматозоиды (имеющие закрученную форму и снабженные двумя и более ресницами) проникают в архегоний через соединительный слой воды, причем возбудителем их движения к архегонию у папоротников и хвощей является яблочная кислота, у плаунов — лимонная. Из оплодотворенной яйцевой клетки развивается бесполое поколение, то есть окончательное взрослое растение, которое размножается только образующимися в определенных местах спорами. Они развиваются в мелких спорангиях, причем каждая материнская клетка дает начало четверке спор. В этот момент происходит редукционное деление. У наших папоротников спорангии почти всегда сидят пучками на нижней стороне листьев; у хвощей они находятся на нижней поверхности особых гвоздевидных чешуй, собранных верхушечным колосом, а у плаунов — в пазухах чешуевидных листьев.
Большинство современных папоротникообразных принадлежит к равноспоровым, то есть образующим споры только одного типа. В прежние геологические эпохи были широко распространены ныне почти вымершие разноспоровые высшие споровые. У них образуются двоякого рода спорангии и двоякого рода споры: более крупные — макроспорангии и макроспоры и более мелкие — микроспорангии и микроспоры (см. XXXI, 135/37 и ХХХII, 335). Особенности разноспоровых позволяют понять, каким образом из типа развития и размножения, свойственного высшим споровым и характеризующегося крайне резко выраженным чередованием поколений, в ходе эволюции возник тип цветковых, у которых чередование поколений кажется совершенно отсутствующим, в действительности же имеется, но сведено до трудно заметных следов. У цветковых половое поколение — заросток — редуцирован до крайних пределов и развивается на бесполом поколении. Этот процесс редукции уже можно проследить у разноспоровых. Их заростки так малы (см. XXXI, 137), что могут почти не выходить наружу из оболочки споры. Соответственно упрощается строение антеридиев и архегониев. При этом наблюдается разделение полов: макроспора дает начало женскому заростку (несущему только архегонии), микроспора — мужскому (несущему только антеридии и подвергающемуся особенно далеко идущему уменьшению и упрощению). Чтобы произошло оплодотворение, необходимо, чтобы ветер перенес микроспоры в ближайшее соседство с макроспорами. У некоторых современных разноспоровых (Selaginella, см. ХХХVIІ, 640) можно наблюдать, что макроспоры начинают образовывать заросток, находясь еще в спорангии на материнском растении. В таких случаях микроспора должна для процесса оплодотворения попасть на макроспорангий и здесь прорасти в заросток. Это и есть исходная форма процесса опыления и оплодотворения, характерного для цветковых растений.
6. Цветковые растения. Наибольшей дифференцировки, как внешней, так и внутренней, и приспособляемости к условиям существования достигают цветковые, или семенные растения (см. цветковые растения). Им удается в сильнейшей степени освободиться от зависимости от избытка воды как в вегетативном состояния, так и в процессе полового размножения. В связи с этим вся картина размножения, сопровождающаяся образованием цветка, по внешности не имеет ничего общего со споровыми растениями. Зачатком нового растения, отделяющимся от материнской особи, у цветковых является уже не микроскопическая клетка, спора, а сложно устроенное и разнообразно вооруженное в борьбе за существование семя, содержащее в себе готовый зародыш. Вместе с тем весь ход индивидуального развития организма приобретает цельность единства без видимого распадения на два неодинаковых по своей структуре и биологическим запросам поколения.
Наиболее близки еще к высшим споровым голосемянные (см. XLV, ч. 3, 176, 178), куда в нашей природе относятся хвойные (см. XLV, ч. 3, 180): наиболее далеко отклоняются от споровых покрытосемянные (см. XLV, ч. 3, 184, сл.), к которым принадлежит громадное большинство современных цветковых. Среди покрытосемянных однодольные отличаются во всех отношениях большей примитивностью своих организационных черт, чем двудольные. Только эти последние проявили наибольшую гибкость, дали наибольшее видовое разнообразие и явились истинными завоевателями современной суши.
Ткани. У цветковых растений тканевая дифференцировка достигает наибольшей типичности, оформленности и в то же время пластичности. Все органы цветковых защищены покровной тканью. Первичной и наиболее распространенной формой ее является кожица, или эпидермис. Она представляет собой прозрачную пленку, состоящую из одного или нескольких слоев клеток, прекрасно приспособленную к регулированию испарения и газового обмена и в то же время не препятствующую прониканию световых лучей к глубже лежащим зеленым клеткам. Клетки кожицы, кроме определенных мест, называемых устьицами (см.), тесно смыкаются друг с другом без межклетных промежутков. Стенки их в зависимости от привычных условий обитания растения в различной степени утолщены. В условиях достаточного обеспечения водой утолщается, и то слабо, только наружная стенка, в условиях более ксерофитных утолщение может распространяться на всю окружность клетки и достигать такой степени, что полость клетки почти исчезает (см. водохранилища у растений и ксерофиты). Самый поверхностный слой наружной стенки клеток кожицы, соприкасающийся с воздухом, так называемая кутикула, приобретает химические и физические свойства, более или менее близкие к пробковому веществу, и усиливает непроницаемость кожицы. Степень развития кутикулы опять-таки бывает весьма разнообразна и может давать мощные кутикулярные слои (см.). Нередко кожица, кроме того, на поверхности выделяет воскообразное вещество (рис. 5), обычно в виде сизого, легко стирающегося налета (например, на плодах слив). У некоторых растений (например, у перуанской «восковой пальмы», Ceroxylon andicola) это выделение достигает размеров целой корки, толщиной в несколько миллиметров, и самый воск находит себе применение в промышленности («растительный воск», см. растительные соки).
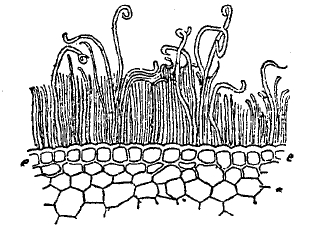
Рис. 5. Кожица (е) с восковыми палочками на поперечном разрезе стебля сахарного тростника.
Особенно часто кожица образует в большем или меньшем изобилии выросты — волоски (рис. 6), чешуйки, бугорки, иногда шипы.
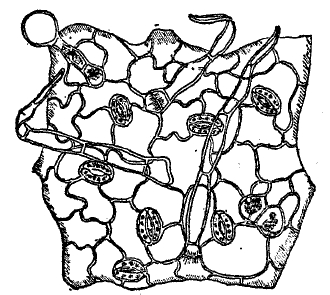
Рис. 6. Кожица с волосками и устьицами.
Форма, строение и биологическая роль волосков (см.) весьма разнообразны. Очень часто волоски рано теряют живое содержимое и, благодаря прониканию в них воздуха, делаются белыми. Очень своеобразный случай представляют жгучие волоски крапивы. Они состоят из длинной, постепенно заостряющейся клетки, сидящей на многоклеточном основании. Кончик волоска отличается большой хрупкостью. Он легко проникает в поры кожи и здесь на верхушке обламывается. Тогда из образовавшегося отверстия в ранку выходит едкое содержимое, вызывающее характерное болезненное ощущение (см. крапивные: ср. железы, XX, 128/29). Клетки кожицы обычно не содержат хлорофилла и богаты водой.
Почти все указанные особенности имеют своей задачей усиление непроницаемости кожицы и локализацию испарения и газового обмена в определенных местах — устьицах (рис. 6 и 7)
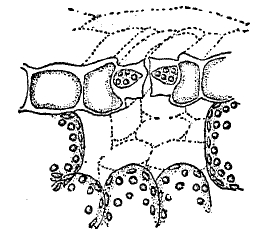
Рис. 7. Устьица в разрезе.
Устьица представляют собой щели, ограниченные двумя замыкающими клетками каждое и обладающие замечательной способностью автоматически закрываться и раскрываться в зависимости от баланса влажности (см. устьица). На долговечных зимующих органах — стеблях и корнях деревянистых растений — кожица заменяется залагающейся под ней более мощной пробковой корой, которая состоит из большого числа слоев отмерших опробковевших клеток. Для газового обмена на местах, соответствующих устьицам, образуются участки рыхлой ткани — чечевички, по межклеточникам которой и могут диффундировать газы (ср. XLIII, 133/34).
Проводящая система у цветковых растений слагается из тончайших длинных трубок двоякого типа. Одни из них — водоносные сосуды — проводят специально воду с содержащимися в ней минеральными солями, другие — ситовидные, или решетчатые трубки, служат для транспортировки сложных органических соединений. В соответствии с различной степенью спешности той и другой задачи, и строение тех и других неодинаково. Водоносные сосуды представляют собой трубки в буквальном смысле слова, получающиеся из продольных рядов клеток путем исчезновения разделяющих их перегородок. Они совершенно лишены живого содержимого. Стенки водоносных сосудов всегда подвергаются процессу одеревенения. При этом для них является характерным неравномерное утолщение с внутренней поверхности в виде правильных спиральных лент (рис. 8), колец, сетки (спиральные, кольчатые, сетчатые сосуды).
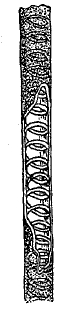
Рис. 8. Сосуд со спиральным утолщением внутри.
Задачей подобного утолщения является противодействие сдавливанию полости трубки окружающей тканью, чередование же утолщенных мест оболочки с тонкими сохраняет возможность легкой боковой отдачи воды, двигающейся по сосудам. Просвет водоносных сосудов незначителен, едва заметный простым глазом, но так как трубки всегда собраны в большом количестве тяжами, то и проводящая способность их соответственно повышается. Сами по себе мертвые, водоносные сосуды всегда сопровождаются обкладками жизнедеятельной древесинной паренхимы (то есть ткани, состоящей из более или менее равномерных изодиаметрических клеток). У голосемянных настоящие водоносные сосуды еще отсутствуют, а имеются лишь сильно вытянутые в длину клетки - трахеиды (см.), отделенные друг от друга на концах косыми перегородками.
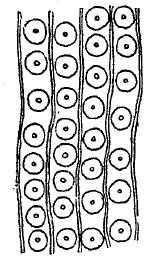
Рис. 9. Трахеиды сосны с окаймленными продушинами.
Проведение по ним воды, конечно, должно быть менее совершенным, чем по настоящим сосудам. Для трахеид хвойных весьма характерны сложно устроенные окаймленные продушины (поры) в оболочках, имеющие под микроскопом при наблюдении с поверхности вид двух концентрических кружков (рис. 9). Их канал посередине шире внутри, чем к обоим концам, и разделен тонкой перегородкой с дискообразным утолщением в центре. Двойной контур продушины слагается из наружного контура сечения самого канала и контура утолщенного диска. Предполагается, что описанное строение окаймленных продушин позволяет им работать наподобие клапанов, причем диск может придавливаться к стенкам канала и закупоривать выход. Ситовидные, или решетчатые трубки (рис. 10) в отличие от трахей сохраняют до конца жизни перегородки между составляющими их вытянутыми в длину клетками, но перегородки эти мелко продырявлены наподобие сита.
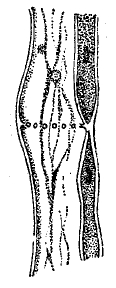
Рис. 10. Решетчатая трубка.
Стенки ситовидных трубок не утолщены и сохраняют до конца живое содержимое, но до самого последнего времени считалось, что ядра в них исчезают. Однако, новейшие исследования Шмидта с применением более точных методов обнаружили присутствие ядер и во вполне развитых ситовидных трубках. По длине ситовидные трубки сопровождаются прилегающими к ним вытянутыми в длину клетками-спутниками с обильным плазматическим содержимым и ядрами. Они являются, быть может, железистыми клетками, выделяющими те или другие вещества, принимающие участие в работе ситовидных трубок. Водоносные сосуды и ситовидные трубки всегда располагаются вместе, образуя сосудисто-волокнистые пучки (рис. 11 и 12), причем часть пучка, состоящая из водоносных сосудов, называется древесиной, или ксилемой (см. древесина), а часть, состоящая из ситовидных трубок, называется лубом, или флоэмой. В виде редкого явления (у тыквенных) луб примыкает с двух сторон к древесине, обычно же имеется только один участок древесины и один участок луба. В состав сосудисто-волокнистого пучка входят и другие клетки: паренхима древесинная и лубяная, затем вытянутые в длину толстостенные механические волокна — лубяные и древесинные (иначе называемые либриформом). Лубяные волокна дают прядильный материал у большинства волокнистых растений (лен, рис. 13; конопля, кендырь, но не хлопчатник!). Сосудисто-волокнистые пучки залагаются в виде островков или кольца вытянутых клеток, называемых прокамбием. У однодольных весь прокамбий исчерпывается сразу на образование пучка. Такой сосудисто-волокнистый пучок уже не обладает способностью к дальнейшему разрастанию (замкнутые сосудисто-волокнистые пучки).
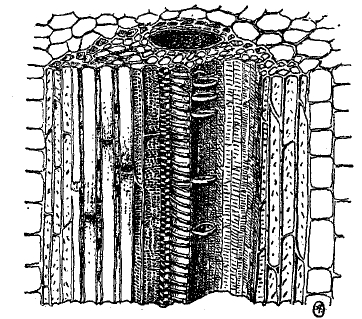
Рис. 11. Сосудисто-волокнистый пучок, разрезанный вдоль.
У двудольных между древесиной и лубом остается прослойка удлиненных эмбриональных клеток — камбия (см.), который затем может в многолетних органах — стеблях, корнях — ежегодно давать все новые и новые отложения элементов древесины и луба (вторичное утолщение).
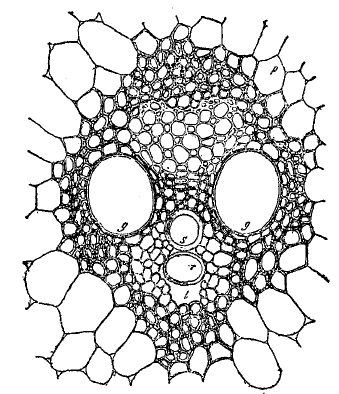
Рис. 12. Сосудисто-волокнистый пучок кукурузы в поперечном сечении; g, g, s, r - сосуды ксилемы, l - межклетный ход, p - тонкостенная паренхимная клетка.
В некоторых семействах (молочайные, маковые, сложноцветные и др.) имеется третий тип проводящих трубок — млечные сосуды (см. сосуды млечные).
Рис. 13. Волокна льна.
Они пронизывают все растения в виде сетки (рис. 14) и не находятся в прямой связи с сосудисто-волокнистыми пучками.
Рис. 14. Сеть млечных сосудов.
Млечные трубки, вероятно, служат в первую очередь для передвижения различных сложных веществ. В пользу этого говорит тот факт, что между степенью развития млечных сосудов и ситовидных трубок наблюдается в известных пределах обратное соотношение. Состав млечного сока отличается сложностью. В нем часто содержится каучук.
Наконец, механическая ткань, служащая для прочности и характеризующаяся, вообще говоря, сильно утолщенными оболочками у цветковых растений, представляет несколько типов. Из них колленхима (рис. 15) является наиболее слабой; ее оболочки утолщаются только в углах или на отдельных стенках, тогда как в остальном остаются тонкостенными. При этом они сохраняют свои целлюлозные свойства. Благодаря указанным особенностям, клетки колленхимы хотя и приобретают некоторую прочность, но сохраняют свою жизнедеятельность.
Рис. 15. Колленхима.
Колленхима (см.) всегда залегает на поверхности органов, под кожицей и содействует укреплению молодых растущих или вообще однолетних органов. Наиболее характерной механической тканью является склеренхима (см.). Она состоит из вытянутых в длину заостренных клеток с сильно утолщенными одеревеневшими стенками (см. стереиды). Третий тип механических клеток — склереиды, или каменистые клетки (см.) — представляет паренхиматические клетки с сильно утолщенными стенками. Они служат для придания прочности частям растения, имеющим более или менее шаровидную форму (например, плодам).
7. Органы цветковых растений. Исходным образованием, с которого начинается развитие новой особи, у цветковых является семя (см. зародыш растения). В семени с самого начала имеется готовый зародыш — крохотный зачаток будущего растения, обладающий уже всеми его частями: стеблем, корнем, листьями. При этом первые шаги его развития обеспечиваются запасом питательных веществ, отлагаемых или в первых листочках зародыша – семядолях (например, у гороха и других бобовых), или в особой ткани, примыкающей к зародышу и называемой эндоспермом (у злаков), реже окружающей его (перисперм). Семя покрыто то более, то менее толстой кожурой, которая хорошо предохраняет его от высыхания и других неблагоприятных воздействий среды. Нередко семена бывают снабжены особыми приспособлениями для содействия их расселению (ветром, животными). Стебель зародыша оканчивается верхушечной почкой, которая растет, постепенно развертывается и дает все новые и новые листья. Все три основных органа, намечающиеся уже у зародыша, характеризуются достаточно постоянными и определенными особенностями, стоящими в связи с их физиологической ролью.
Лист. Лист (рис. 16), как орган ассимиляции (см. фотосинтез) и испарения (см. транспирация), состоит обычно из тонкой, но развитой в плоскости пластинки и стебельчатой части, поддерживающей ее, черешка, хотя бывают листья и сидячие, то есть прикрепляющиеся непосредственно пластинкой к стеблю. Черешок нужно рассматривать как специальное приспособление для регулирования наиболее выгодной световой установки пластинки. Благодаря изгибам черешка, верхняя сторона пластинки всегда обращена к свету.
Рис. 16. Лист с прилистниками.
При основании листа иногда бывают мелкие листочки — прилистники. У некоторых растений (особенно часто у однодольных: злаков, осок и др.) основание листа расширяется и образует вокруг стебля влагалище. Размеры листьев колеблются в чрезвычайно широких пределах; точно также и форма их представляет громадное разнообразие. Листья пальм и некоторых других тропических растений так велики, что одним листом можно накрыть целую хижину; с другой стороны, у растений-обитателей пустынь сплошь и рядом листья низводятся до незначительных чешуй. Причина таких колебаний кроется частью во внешних условиях, частью во внутренних организационных свойствах самого растения. Чем более условия существования благоприятны в смысле обеспечения водой, тем большая развивается поверхность листьев. Обратно, при неблагоприятном балансе влажности и листовая поверхность уменьшается иногда до полного исчезновения (у кактусов).
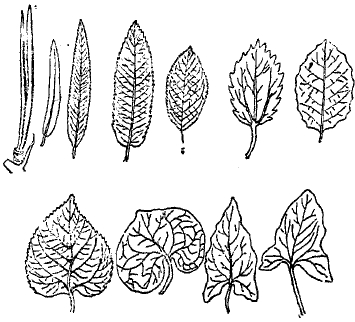
Рис. 17. Различные формы листьев.
Но вместе с тем, например, злаки никогда не дают больших широких листьев; форма листа у них колеблется лишь в пределах от узколинейной до широко-ланцетной. Форма листьев определяется, с одной стороны, общим очертанием краев и с другой - степенью надрезанности.
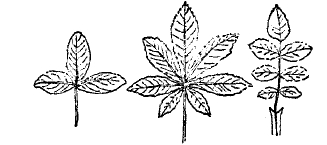
Рис. 18. Тройчатый, пальчатый и перистосложный лист.
По очертанию листья бывают: сердцевидные, круглые, эллиптические, продолговатые, яйцевидные, ланцетные, линейные и др. (рис. 17). По степени надрезанности листья могут быть: совершенно цельнокройными, зубчатыми, лопастными, раздельными, рассеченными и, наконец, сложными (рис. 18). Эти последние, в отличие от простых листьев, несут на общем черешке по нескольку или по многу совершенно самостоятельных пластинок, или листочков. В более типических случаях (например, у мотыльковых) листочки сложного листа снабжены при основании особым сочленением, позволяющим им совершать изменения положения (например, дневное, ночное положение). Листочки сложного листа, как и надрезы листьев вообще, могут располагаться дланевидно, или пальчато (конопля, лютик, клен), то есть сидеть в одной точке на верхушке черешка, или же перисто (например, акация), с парами листочков, отодвинутыми друг от друга по длине черешка. Форма листьев очень постоянна для целых семейств; так, у гвоздичных листья всегда цельные, цельнокрайные, от линейных до эллиптических; у зонтичных листья глубоко и повторно рассеченные; у мотыльковых — сложные. С другой стороны, приспособление к сходным условиям существования приводит к сходству листьев у самых различных растений. Так, листья водяных растений (см.), плавающие на поверхности воды, имеют ясную наклонность к цельнокрайности и к широко эллиптической, до почковидной, форме. Наоборот, погруженные в воду листья у самых разнообразных растений узколинейны или же глубоко рассечены на узколинейные (до волосовидных) доли.
Рис. 19. Дугонервный лист ландыша.
У некоторых растений (например, у жерухи земноводной) подводные и воздушные листья связаны переходами в зависимости от степени погруженности в воду. Внешние условия, однако, как показывают опыты, в таких случаях не непосредственно создают ту или другую форму листа, а лишь дают толчок к скрытой в организме формообразовательной реакции. Пластинка листа пронизывается тяжами, нервами, или жилками, выступающими над поверхностью с нижней стороны листа. Они представляют собой систему сосудисто-волокнистых пучков, служащих для снабжения листьев водой и минеральными солями и вместе с тем играющих рель каркаса, механически поддерживая и расправляя тонкую пластинку. В связи с этим в жилках всегда имеется механическая ткань, в частности, в выступающих над поверхностью частях их залегает колленхима.
Характер пробегания листовых нервов (нервация), за немногими исключениями, сильно отличается у однодольных от двудольных. В первом случае листья бывают параллельно-нервными и дугонервными (рис. 19), то есть жилки пробегают от основания листа к его верхушке или параллельно, или дугообразно, причем боковые соединения между ними (анастомозы) слабо выражены. Во втором случае господствует тип сетчато-нервных листьев, при котором нерватура представляет более или менее густую сетку (рис. 20).

Рис. 20. Дланевидно-лопастный лист клена.
При этом в перисто-нервных листьях имеется одна главная, более сильная жилка, или главный нерв, остальные же отходят от этого последнего по его длине; в пальчато-, или дланевидно-нервных листьях главных нервов несколько, и они все расходятся веерообразно от основания пластинки. В листьях типа однодольных перерезка тех или других жилок выводит из снабжения водой весь дальнейший участок пластинки. В сетчато-нервных листьях подобная перерезка не оказывает заметного влияния, так как легко устанавливаются обходные пути.
Во внутреннем строении пластинки листа наземных цветковых в условиях среднеевропейского климата микроскоп обнаруживает следующие характерные черты строения (рис. 21). Сверху и снизу лист покрыт кожицей. Устьица обычно сосредоточены в кожице нижней стороны. Под верхней кожицей залегает один-два слоя клеток столбчатой, или палисадной ткани (см. столбчатая ткань), заметно вытянутых в длину и стоящих перпендикулярно к поверхности листа. Они богаты хлорофилльными зернами. Под столбчатой тканью залегает губчатая ткань (см.), клетки которой имеют обыкновенно извилистую форму и смыкаются друг с другом, оставляя большие межклетные ходы. Система этих последних сообщается непосредственно с устьицами, под которыми залегает особенно крупный межклетник (дыхательная полость). Содержание хлорофилльных зерен в губчатой ткани значительно уступает палисадной ткани. Столбчатая ткань по своему положению, наиболее выгодному в световом отношении, вместе с тем является тканью листа, наиболее интенсивно ассимилирующей.
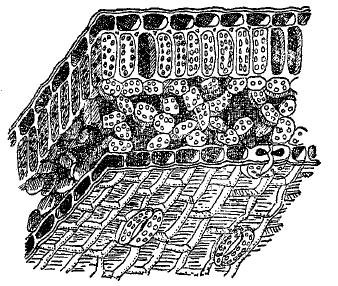
Рис. 21. Строение листовой пластинки в разрезе.
Ассимиляционная роль губчатой ткани, уже по своему более глубокому положению получающей менее света, отступает на задний план. Благодаря развитию межклетников, губчатая ткань допускает легкий доступ к палисадным клеткам углекислоты, поступающей из воздуха через устьица, и обратную отдачу кислорода. В природе встречаются самые разнообразные уклонения от описанного типа анатомического строения листа. В этом отношении лист отличается чрезвычайной пластичностью и может служить прекрасным показателем экологических условий. Так, у растений, живущих в сильном затенении (например, под пологом леса), палисадная ткань, по своему строению рассчитанная на глубокое проникание световых лучей, слабо выражена; клетки ее делаются низкими и широкими, не резко отличаясь от губчатой ткани. Дифференцировка листа еще более понижается в подводных листьях цветковых. Наоборот, в условиях обитания, характеризующихся повышенным освещением, наблюдается мощное развитие палисадной ткани. У ксерофитов при этом палисадная ткань может развиваться и на нижней поверхности листа, губчатая же паренхима отодвигается в середину листа или почти отсутствует. У листьев водяных растений, плавающих на поверхности воды и находящихся также в условиях полного освещения, палисадная ткань или ряды палисадовиднорасположенных коротких клеток образуют значительную толщу, но между ними образуются большие межклетные пространства. Даже на одном и том же растении, особенно на деревьях, можно наблюдать большие или меньшие различия в анатомическом строения листа, в зависимости от положения листа на поверхности кроны или в глубине ее (световой и теневой лист). Нижняя сторона листа, на которой находятся устьица, очень часто бывает защищена вспомогательными образованиями, облегчающими регулирующую роль устьиц: густым волосистым опушением, восковым налетом.
В нашей природе господствуют листья, сбрасываемые на зиму (см. листопад). В климатах с безморозными зимами получают широкое распространение вечнозеленые листья. Они отличаются большей плотностью, кожистостью и глянцевитостью, зависящей от сильно развитой кутикулы. Вечнозеленые листья в действительности также время от времени заменяются новыми, но это сбрасывание происходит не каждый год и не одновременно, поэтому и мало заметно. У нас к вечнозеленым породам принадлежит только большинство хвойных.
Места прикрепления листа к стеблю получили название узлов, разделяющие же их участки стебля — междоузлий. В распределении листьев на стебле (листорасположении) наблюдаются закономерности, весьма постоянные для целых систематических групп. Наиболее обычный тип листорасположения — спиральный, или очередной, когда на одной высоте от стебля отходит каждый раз всего один лист, причем все листья сидят по ходу очень правильно идущей спирали. Ее можно начертить на поверхности стебля, соединяя последовательно непрерывной линией листья от основания одного листа до основания ближайшего следующего, идя все время в одном направлении. Оказывается, что углы, отделяющие два смежных листа, отличаются большим постоянством. Величину этого угла принято выражать не в градусах, а в частях окружности — дробью, называемой формулой листорасположения. Чтобы получить ее, берется число оборотов, которое описывает спираль от одного листа до ближайшего следующего, находящегося как раз на одной и той же продольной линии (ортостихе), и делится на число этих последних, так как листья в пределах спирали расположены продольными рядами. У злаков, например, листья располагаются в 2 ряда, и спираль делает всего один оборот; поэтому формула листорасположения 1/2; у осок — 1/3; у подсолнечника (рис. 22) — 1/5 и т. д. Замечательно, что наиболее распространенные формулы листорасположения представляют собой первые приближения непрерывной дроби
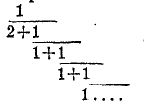
Рис. 22. Схема расположения листьев на стебле подсолнечника.
Механизм происхождения подобной математической закономерности нельзя считать вполне выясненным. По взгляду, обоснованному Швенденером, в основании его лежат чисто механические взаимоотношения, создающиеся между полушаровидными бугорками - зачатками листьев на тесной поверхности верхушечного конуса нарастания, но под это объяснение могут быть подведены далеко не все случая. Кроме очередного листорасположения, встречаются: супротивное (по два листа на одной высоте) и кольчатое, или мутовчатое (по 3 и больше листа на одной высоте).
Листья закладываются на поверхности стебля в точке роста в виде бугорков. Дифференцировка листа идет таким образом, что растет основание, верхушка же является наиболее старой частью. Кроме верхушечной почки, в план строения листостебельного побега всегда входят пазушные почки, находящиеся в углах, образуемых основаниями листьев и стеблем. Каждая пазушная почка может дать начало боковой ветке, и ветвление стебля должно было бы быть полным повторением листорасположения. В действительности большая или меньшая часть почек с определенной правильностью не развивается дальше, а замирает. В результате получается значительное разнообразие ветвления, а вместе с тем и внешнего вида растения (см. XII, 161/62). Кроме нормальных — верхушечной и пазушных почек, могут развиваться на разных местах придаточные почки, возникающие главным образом в связи с поранением (см. почка).
Как ближайшие к основанию, так и ближайшие к верхушке побега листья обыкновенно более или менее отличаются от средних, наиболее типично выраженных. Из низовых листьев особенно выделяются семядоли, которые в большинстве случаев отличаются большей простотой формы, меньшей величиной и нередко отсутствием до конца зеленой окраски.
Рис. 23. Листообразные черешки у австралийской акации.
К низовым листьям принадлежат также чешуи почек, которые представляют собой сильно уменьшенные и измененные листья или их части. Особенно велико уклонение от первоначальной формы у зимующих почек древесных пород, которые закладываются в один вегетационный период, а развиваются только через зиму. У них наружные чешуи играют защитную роль от высыхания и разных колебаний температуры, в связи с чем они изменяются в сторону большей плотности и кожистости и покрываются нередко волосками или смолистыми выделениями. Из верховых листьев наиболее сильно уклоняются разнообразные листья, входящие в состав цветов и соцветий: здесь они могут совершенно утрачивать свою обычную форму и окраску (метаморфоз листа).
Иногда листья берут на себя особые задачи и соответственно претерпевают изменения. Так, листья могут быть превращены в усики, служащие для обхватывания других растений и поддерживания слабого стебля лазящих растений, в колючки и проч. В некоторых случаях функция пластинки переходит на черешок: этот последний тогда пластинчато расширяется (филлодий), собственно же пластинка недоразвивается (у австралийских акаций; рис. 23).
Стебель. На стебель выпадает роль служить поддерживающим механическим остовом для всей системы листьев, образовать эти последние, и заменять отработавшие новыми, способствовать их наиболее выгодному световому положению и служить связующей системой для проведения веществ из корней в листья и обратно. Стебель вместе с тем является наиболее долговечным органом растения, берущим на себя у многолетних растений задачу сохранения жизнедеятельности в течение зимнего или сухого летнего периода. В связи с указанной ролью стебель вытягивается в одном направлении и уже с момента прорастания зародыша в своем росте упорно направляется кверху вследствие отрицательного геотропизма (см. тропизмы).
Рис. 24. Точка роста стебля с зачатками листьев.
Кроме геотропической чувствительности, стебель обыкновенно обладает положительным фототропизмом. Он растет своей верхушкой, на которой образуются все новые и новые листья в виде почки (рис. 24), но кроме того нередко имеет место вставочный (интеркалярный) рост на протяжении междоузлия. В анатомическом строении стебля на передний план выступает проводящая система. Широко развит процесс одеревенения оболочек.
Внешняя форма стебля сравнительно однообразна. На поперечном сечении стебли бывают чаще всего круглыми, но могут быть треугольными, четырехугольными, сплюснутыми и проч. У вьющихся и лазящих растений (см. XXVI, 380/82), или лиан, стебель сам по себе слаб и не держится прямо, но он взбирается вверх и выносит к свету листовую массу, благодаря обладанию особыми хватательными органами — усиками, или же благодаря обвиванию вокруг других более прочных растений. И усики, и стебли вьющихся растений совершают своей верхушкой движения и обладают вместе с тем повышенной чувствительностью к соприкосновению (тигмотропизм). Лежачие и ползучие стебли совсем не поднимаются над землей. Во многих случаях определенные междоузлия стебля остаются укороченными, незаметными для глаза. Таким путем получаются прикорневые розетки листьев, круги листочков околоцветника в цветах. Стебли со слабым одеревенением называются травянистыми и обыкновенно сохраняют свою жизнедеятельность только один вегетационный период. Напротив того, деревянистые стебли деревьев и кустарников (кустарник от дерева отличается ветвлением ствола от самого основания) могут существовать очень продолжительное время, до нескольких тысяч лет (баобаб, мамонтово дерево и др.). Но и растения травянистые могут иметь различную продолжительность жизни. В этом отношении различают: однолетники, то есть растения, успевающие в течение одного вегетационного периода вполне развиться и принести семена, после чего они погибают; двулетники — растения, требующие для своего полного цикла развития двух вегетационных периодов; в первый год они обыкновенно не образуют развитого стебля, а лишь прикорневую розетку листьев, отлагая запасы питательных веществ в корнях, стебель же, цветы и плоды развиваются на второй год, после чего растение погибает (морковь, свекла). Посредине между однолетниками и двулетниками стоят озимые растения. Они начинают развиваться с осени; с первыми листьями подходят под снег и кончают свое развитие на следующий год. Будучи высеяны весной, они все же в первый вегетационный период не развиваются дальше. Наконец, существует большое число многолетних трав. Их жизнедеятельность, несмотря на ежегодное отмирание надземных частей, сохраняется в течение многих лет, благодаря образованию подземных стеблей. Подземные стебли представляют три основных типа: корневище (см.), клубень (см. клубни) и луковицу. Корневище по внешности напоминает корень. Многолетние травы с различно развитыми корневищами образуют основу луга целинной степи. Корневища могут быть более или менее ползучими (рис. 25) и давать побеги, отстоящие друг от друга на значительное расстояние (рыхло-кустистые многолетники), или же быть сильно укороченными и ветвиться в тесном пространстве (густо-кустистые многолетники). Клубень представляет собой укороченное и сильно утолщенное корневище (например, клубни картофеля: они образуются не на корнях, как можно думать при поверхностном взгляде, а на подземных ветвях стебля). У луковицы стебель остается очень коротким и ширококоническим (донце) и несет мясистые чешуйчатые листья, богатые запасными питательными веществами.
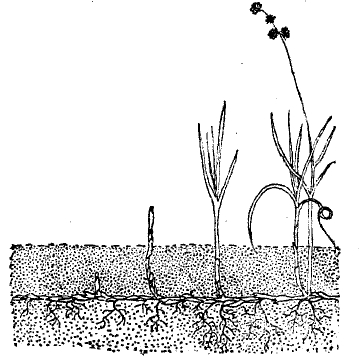
Рис. 25. Ползучее корневище осоки.
Надземные побеги ежегодно образуются или из верхушечной почки, или из пазушных.
Однолетние стебли на поверхности покрыты обыкновенной кожицей, под которой залегает на некоторую глубину зеленая паренхима, и таким образом могут в известной степени ассимилировать. У резко выраженных сухолюбов листья могут совсем не развиваться, и тогда стебель всецело берет на себя роль ассимилирующего органа. Иногда при этом ветви стебля принимают и листообразную форму (кладодии, см.; у Ruscus, рис. 26). На поперечном разрезе однолетних стеблей под залегающей на поверхности кожицей бросается в глаза заполняющая его масса основной паренхимы, содержащей хлорофилл только в слоях, ближайших к поверхности. У двудольных под кожицей залегает обычно колленхина, служащая для укрепления стебля.

Рис. 26. Листообразные ветви у Ruscus.
В основной паренхиме залегают более или менее многочисленные сосудисто-волокнистые пучки. Они в стеблях всегда обращены своей лубяной частью к периферии стебля, а древесиной к центру.
Рис. 27. Распределение сосудисто-волокнистых пучков на поперечном разрезе стебля однодольного растения.
В расположении сосудисто-волокнистых пучков различаются два типа: однодольных и двудольных. У первых сосудисто-волокнистые пучки разбросаны без определенной правильности по всему поперечнику стебля (рис. 27), у вторых они располагаются кольцом, оставаясь обособленными друг от друга в виде отдельных островков или же сливаясь вместе в непрерывный круг. Беспорядочное расположение сосудисто-волокнистых пучков у однодольных внутренне ясно связано с отсутствием у них камбия и вторичного утолщения, тогда как правильное кольцеобразное расположение пучков сделало возможным отложение все новых и новых слоев древесины и луба. Процесс вторичного утолщения стебля у цветковых открыл широкий простор для нарастания массы отдельных индивидуумов. В связи с ним приходится говорить отдельно о первичном и вторичном строении стебля.
В первичном строении стебля двудольных та часть основной паренхимы, которая лежит внутри кольца, образуемого пучками, называется сердцевиной, а радиальные полоски паренхимы, находящиеся в промежутках между отдельными пучками, — первичными сердцевинными лучами (см. сердцевинные лучи). Кнаружи от сосудисто-волокнистых пучков лежит первичная кора (см. кора). По направлению внутрь стебля первичная кора не доходит до самых сосудисто-волокнистых пучков. Внутреннюю ее границу составляет слой клеток, называемый эндодермой. Они выделяются содержанием крахмальных зерен, отличающихся здесь легкой смещаемостью: скопления зерен в клетках эндодермы играют роль простейших органов чувства равновесия (статоцистов). Между эндодермой, принадлежащей еще первичной коре, и сосудисто-волокнистыми пучками лежит то более толстый, то более тонкий кольцеобразный слой клеток перицикла, нередко сильно утолщенных. Перицикл вместе со всем, лежащим внутри его, образует центральный цилиндр.
При вторичном утолщении сосудисто-волокнистые пучки, если они образовали до того отдельные островки, сливаются в одно кольцо, путем возникновения из межпучкового камбия (см. камбий). В дальнейшем клетки камбиального кольца начинают правильно периодически делиться и отлагать новые клетки, как по направлению внутрь ствола — к древесине, так и к поверхности — лубу. Первые постепенно превращаются соответственно в водоносные сосуды и другие элементы древесины, вторые — в ситовидные трубки и другие элементы луба. Таким образом, образуются вторичная древесина (см. древесина) и вторичный луб. Благодаря отложению все новых и новых слоев древесины, ствол непрерывно нарастает в толщину. Что касается луба, то нарастание его идет совнутри и имеет неизбежным последствием пассивное растяжение лежащих кнаружи более старых тканей, которые, в конце концов, от сдавливания отмирают и постепенно слущиваются. Раньше всего этой судьбе подвергается кожица, а также нередко первичная кора. На местах сбрасывания старой ткани образуется пробковая ткань вследствие возникновения особого деятельного слоя клеток - пробкового камбия (см. феллоген). Таким образом, толща многолетних стволов состоит главным образом из вторичной древесины; вторичный же луб образует вокруг нее лишь тонкий слой, составляя вместе с пробковой тканью вторичную кору (см. кора). Благодаря вторичному утолщению, стволы деревьев на поперечном разрезе (рис. 28) представляют характерную концентрическую слоистость, причем нормально каждый слой соответствует одному году (см. древесина).

Рис. 28. Поперечный разрез ствола двудольного растения.
Так как ширина слоя, отлагаемого камбием, зависит от условий питания, в каких находилось дерево, то, изучая характер годичных колец, можно получить представление об условиях освещения и влажности данного года или вообще местообитания. На поперечных разрезах древесных стволов видны прямые линии, радиально расходящиеся в древесине от внутренней части ее до самой поверхности. Они известны под именем вторичных сердцевинных лучей (см. сердцевинные лучи).
Таким образом, ствол дерева превращается, в конце концов, как бы в одну сплошную проводящую воду систему мельчайших трубок древесины, обтянутых на поверхности тонким слоем трубок луба, несущих в обратном направлении сложные органические вещества и облеченных все вместе в непроницаемый футляр пробки. Однако, в действительности не вся масса древесины сохраняет свое активное участие в проведении воды до конца жизни растения. Центральная, старая масса сосудов с течением времени претерпевает различные видоизменения. У одних пород в ней отлагаются разнообразные вещества, придающие этой части ствола нередко особенную плотность. Ее называют ядром, остальную же жизнедеятельную часть ствола — заболонью. Ядро нередко бывает окрашено в различные цвета (черный, желтый, красный и др.) и особенно ценится в технике. У других пород образования ядра не происходит, и старая древесина легко подвергается загниванию, сопровождающемуся образованием дупл (см. древесина).
Корень. Корень (см.), как орган всасывания воды из почвы, в типических случаях имеет и глубоко приспособленное строение.
Рис. 29. Корневые волоски на корешке всхода.
Являясь непосредственным продолжением стебля зародыша (рис. 29), главный корень отличается резко выраженным положительным геотропизмом и настойчиво стремится расти вниз.
Рис. 30. Часть поперечного разреза корня с корневыми волосками (под микроскопом).
Корень всегда имеет цилиндрическую форму, ровную поверхность и на верхушке приострен. Все эти особенности явно стоят в связи с роющей деятельностью корней. Нарастание корня происходит его верхушкой (см. инициала и конус нарастания), но точка роста корня прикрыта особыми защищающими ее слоями клеток, образующими корневой колпачок, или чехлик (см.). Клетки чехлика с поверхности отмирают и слущиваются, а совнутри заменяются новыми. Характернейшей особенностью корня являются корневые волоски (рис. 30). Они представляют собой одноклеточные выросты клеток поверхностного слоя корня и развиваются в большом количестве, начинаясь на некотором расстоянии от верхушки корня. С другой стороны, на старых частях корня волоски отмирают. Благодаря корневым волоскам всасывающая поверхность корней чрезвычайно увеличивается. Кроме того, корневые волоски входят в очень тесное соприкосновение с частицами почвы и таким образом становятся с ними в тесные осмотические соотношения. Однодольные и двудольные по устройству корневой системы представляют характерные различия. У первых главный корень рано отмирает и заменяется более или менее многочисленными придаточными корнями, образующимися от стебля. У вторых главный корень ветвится и дает начало боковым корням.
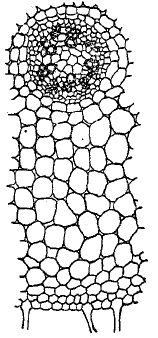
Рис. 31. Микроскопическое строение корня (поперечный разрез).
Анатомическое строение корня (рис. 31), хотя и являющегося непосредственным продолжением стебля, представляет характерные особенности. В корнях сосудисто-волокнистые пучки стянуты к центру, так что сердцевина может совсем отсутствовать, кора же, наоборот, занимает большую часть поперечника корня. При этом участки древесины и луба сосудисто-волокнистых пучков изменяют свое взаимное положение, поворачиваясь на 90° и располагаясь поочередно по радиусам. Эндодерма в корнях сильнее выражена и, благодаря утолщению своих оболочек и частью одеревенению их, частью опробковению, превращается в настоящий внутренний футляр, изолирующий центральный цилиндр. Однако, в этом футляре сохраняются отдельные тонкостенные жизнедеятельные клетки, называемые пропускными, через которые и происходит сообщение коры с центральным цилиндром. Между эндодермой и сосудистой частью цилиндра залегает прослойка перицикла. Замечательно, что в клетках перицикла закладываются боковые корни против древесинных участков и затем прорывают кору (рис. 32). Этот способ заложения, совершенно отличный от заложения ветвей стебля, позволяет сосудистой системе боковых корней непосредственно примыкать к сосудистой системе главного корня. Вместе с тем в связи с указанным способом заложения боковые корни располагаются на главном корне продольными рядами, соответствующими ходу древесинных участков в центральном цилиндре корня. При описываемом строении корня вода с минеральными солями осмотическим путем всасывается корневыми волосками, затем передвигается по клеткам коры до эндодермы, проходит через пропускные клетки и поступает почти непосредственно в водоносные сосуды.
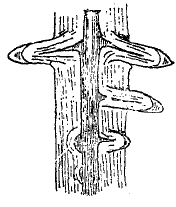
Рис. 32. Способ возникновения боковых корешков (в глубине).
Отсюда делается понятным смысл различного взаимного расположения древесины и луба в стеблях и в корнях, так как при положении луба кнаружи от древесины он стоял бы на дороге поступления воды в сосуды. В старых корнях деревьев первичное строение корня подвергается глубокому изменению вследствие процесса вторичного утолщения. В связи с последним появляется извилистое кольцо камбия, проходящее таким образом, что все участки луба оказываются лежащими кнаружи от него, а древесины — к внутри. В результате отложения новых слоев древесины и луба строение корня делается с трудом отличимым от строения стебля. Корни могут служить местом отложения запасных питательных веществ. В таких случаях они утолщаются конусообразно или в виде клубней и шишек. У тропических орхидных и ароидных наблюдаются воздушные корни (см.).
Чрезвычайная пластичность листьев, наличность корневой системы, способной энергично всасывать воду из глубоких слоев почвы и подавать ее в надземные органы, приспособленность стебля к быстрой подаче воды, все это взятое вместе сделало возможным для цветковых растений существование при самых разнообразных условиях влажности. Среди них, с одной стороны, имеются чисто водные обитатели, а с другой — многочисленные жители самых безводных пустынь, в которых многими месяцами не выпадает ни капли воды. В зависимости от большого разнообразия климатических и почвенных условий на поверхности земного шара, и мир растений представляет значительное число биологических типов, из которых каждый отмечается глубокими приспособлениями к определенной комбинации условий существования (см. фитогеография).
8. Размножение у цветковых растений. Еще более глубокое преобразование претерпел у цветковых способ размножения в результате эволюции в сторону освобождения от зависимости от воды. Вегетативное размножение (см. VIII, 94), по крайней мере, у покрытосемянных, имеет всеобщее распространение, но никогда не происходит путем образования одноклеточных зачатков, аналогичных спорам. Для него служат непосредственно части растения (побеги, корневища, луковички и пр.), которые, с одной стороны, сами достаточно защищены от высыхания, а с другой — долго остаются в связи с материнским растением.
Особенно замечательны изменения в половом процессе цветковых растений. Для обеспечения его возникает сложный орган — цветок. Цветы голосемянных устроены еще очень просто и отчетливо обнаруживают свое происхождение от высших споровых. У них можно проследить, с одной стороны, возникновение при оплодотворении характерного для цветковых растений образования пыльцевой трубочки, а с другой — утрату мужскими клетками подвижности (см. XLV, ч. 3, 176/78). У покрытосемянных строение цветка и картина оплодотворения достигает наибольшей выраженности, причем опять-таки, как и в строении вегетативных органов, однодольные стоят на более ранней ступени, характеризующейся меньшей пластичностью, чем двудольные. Только у двудольных, например, околоцветник приобретает вполне законченное строение из чашечки и венчика. При этом чашечка играет роль защитного образования во время формирования цветка, венчик же — роль привлекающего насекомых аппарата. В цветах однодольных по большей части одни и те же листочки простого околоцветника выполняют ту и другую роль. Точно так же и численные отношения в цветах однодольных гораздо проще, чем у двудольных (см. XLV, ч. 3, 185/204).
Половой процесс у цветковых растений совершенно освобождается от необходимости присутствия воды, без которого невозможно соединение половых клеток у высших споровых. Носительница мужских гамет — пыльца (см.), или цветень, хорошо защищена оболочкой от высыхания и может переноситься через большое воздушное расстояние на аппарат, скрывающий в себе яйцевую клетку — пестик.

Рис. 33. Цветение лесного орешника.
Замечательны те приспособления, при помощи которых осуществляется этот перенос. Они сравнительно просты и однообразны при опылении ветром. Цветы анемофимных, или ветроопыляемых растений характеризуются следующими особенностями (рис. 33): обильным развитием чрезвычайно сыпучей цветени; слабым развитием или полным отсутствием околоцветника, причем яркоокрашенных листочков не бывает; сильным развитием рыльцевой поверхности (см. XII, 174). Анемофилия очень часто сопровождается раздельнополостью (цветы однодомные или двудомные). В тычинковых цветах имеются различные приспособления для облегчения вытряхивания пыльцы при движениях ветра (чрезвычайно тонкие эластические тычиночные нити у злаков, гибкие стержни у соцветий, как, например, у наших деревьев, снабженных сережками).
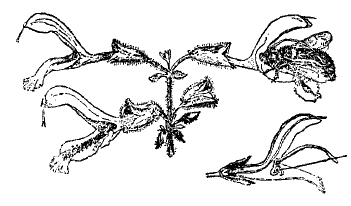
Рис. 34. Механизм опыления у шалфея.
При энтомофилии (опылении насекомыми) общими признаками являются: присутствие привлекающего аппарата в виде ярких лепестков или запаха; образование в цветах продуктов, собираемых насекомыми, в виде меда или, по крайней мере, избыточной цветени; липкость цветени, облегчающая приставание к телу насекомого: обоеполость, связанная с приспособлениями, затрудняющими самооплодотворение (главным образом неодновременное созревание тычинок и пестика); и, наконец, такое устройство и расположение частей цветка, при котором насекомое, посещая этот последний, невольно является переносчиком пыльцы из одного цветка на рыльце другого. В деталях относящиеся сюда приспособления чрезвычайно разнообразны и сплошь и рядом поражают своей замысловатостью (ср. XLV, ч. 3, 196/99).
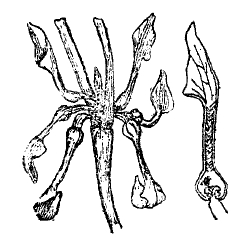
Рис. 35. Цветы кирказона.
У шалфея (рис. 34), например, тычинки устроены наподобие подвижного неравноплечего рычага. Когда насекомое пробирается к меду, оно толкает вперед стоящее на пути короткое плечо рычага, в результате чего длинное, несущее пыльник, ударяет его по спинке и осыпает цветенью. В других, более старых цветах тем же местом спинки насекомое задевает за зрелое рыльце. Особенно оригинально (рис. 35) опыление цветов обыкновенного кирказона (см. кирказоновые). У тропической орхидеи Coryanthes нектар скапливается в большом количестве в особом вместилище наподобие ведра. Насекомые, садясь на край его, легко сваливаются в жидкость и начинают выкарабкиваться из нее. В конце концов, они находят лазейку, через которую и выползают, но она устроена таким образом, что, двигаясь по ней, насекомое обязательно задевает или за пыльник, или за рыльце.
За опылением цветка следует второй акт — оплодотворение. Он начинается с проростков цветени (рис. 36). Под влиянием жидкости, выделяемой рыльцем, пыльцевое зернышко выпускает трубочку, которая, удлиняясь, внедряется в ткань рыльца и идет дальше по столбику. Оболочка пыльцы состоит из двух слоев: наружного — экзины, и внутреннего — интины. В экзине с самого начала имеются особые тонкие места, или окошечки, через которые при прорастании и продавливается интина.
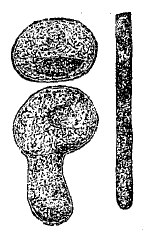
Рис. 36. Образование пыльцевой трубки.
Прорастание цветени вызвать искусственно и вне рыльца, если положить ее в раствор сахара. Пыльцевая трубочка растет до тех пор, пока не достигнет семяпочек. Расстояние, которое ей приходится пройти, иногда бывает весьма значительной длины, достигая в отдельных случаях свыше 10 см. В столбике имеется уже заранее намеченный путь, в виде серединного канала или тяжей особых рыхлых приводящих клеток. В завязи пыльцевая трубочка продолжает свой путь обыкновенно вдоль стенки по имеющимся здесь полоскам такой же проводящей ткани.
Семяпочка, являющаяся носительницей женской, яйцевой клетки и скрытая у однодольных и двудольных внутри завязи (рис. 37), состоит в зрелом виде из центрального округлого или эллипсоидального многоклеточного тела — ядра и облекающих его одного или двух многоклеточных же покровов. Последние оставляют свободным только узкий канал на верхушке семяпочки — семявход. Основание семяпочки — халаза - переходит в семяножку, которая прикрепляется в определенных местах завязи, называемых плацентой. От халязы отходят покровы.
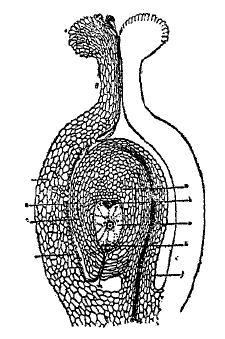
Рис. 37. Строение пестика (с одной обратной семяпочкой).
Чаще всего семяножка так изгибается, что семяпочка обращена к плаценте, а халаза в противоположную сторону (обратная семяпочка). При этом семяпочка частью срастается с поверхностью прилегающей к ней семяножки, образуя семенной рубчик, след которого остается и на семенах. Реже встречается согнутая семяпочка при прямой семяножке или же прямая семяпочка с семявходом, обращенным от плаценты.
Ядро семяпочки скрывает в себе большую пузырькообразную клетку, зародышевый мешок (рис. 38), располагающуюся под самым семявходом. Зародышевый мешок залагается непосредственно под кожицей ядра, почти всегда в виде одиночной крупной клетки.
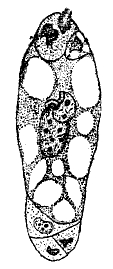
Рис. 38. Зародышевый мешок (выделенный из семяпочки).
В одних случаях в дальнейшем эта клетка непосредственно превращается в мешок, в других, более распространенных, она сначала делится на несколько расположенных продольным рядом клеток, из которых в дальнейшем одна начинает разрастаться и вытеснять остальные. Первичное ядро зародышевого мешка подвергается нескольким повторным делениям. Оно сначала делится на два, которые отодвигаются к двум противоположным концам мешка. Затем каждое из этих дочерних ядер путем двух повторных делений дает начало четырем ядрам (лежащим, следовательно, двумя группами). В дальнейшем от той и другой группы отходит по ядру к середине мешка и сливается, в конце концов, в одно вторичное ядро зародышевого мешка. Остальные ядра, вследствие концентрирования вокруг них протоплазмы, превращаются в голые клетки, лежащие тесно рядом по 3 двумя группами. Из них в той группе, которая лежит у верхушки зародышевого мешка (другими словами, вблизи семявхода), одна клетка представляет собой яйцевую, две же других получили название вспомогательных, или синергид. Клетки же противоположной группы носят название антипод.
Клетки цветени покрытосемянных в начале развития имеют одно ядро, но к моменту окончательного созревания последнее делится на два. Одно из них получает название вегетативного, другое — генеративного. По мере роста пыльцевой трубочки содержимое пыльцы перемещается в конец последней. Вместе е тем, раньше или позже, генеративное ядро делится на два ядра, — спермии, которые по своей роли соответствуют сперматозоидам высших споровых, но лишены ресничек и самостоятельной подвижности. После того как конец пыльцевой трубочки проникнет в семявход и придет в соприкосновение с зародышевым мешком, оболочка его растворяется, и оплодотворяющие ядра выскальзывают в зародышевый мешок. Здесь они расходятся. Одно из них направляется к яйцевой клетке и сливается с ней. Этот процесс слияния есть оплодотворение и дает толчок к развитию яйцевой клетки в зародыш (см. зародыш растения). Другое ядро направляется к вторичному ядру зародышевого мешка и сливается с ним. Это второе слияние дает также толчок к клеточным делениям и приводит к образованию более или менее развитой ткани — эндосперма, служащей местом отложения в таких случаях запасных питательных веществ для зародыша.
Описываемая картина двойного оплодотворения, характерная для покрытосемянных, установлена впервые С. Г. Навашиным. Смысл ее и отношение к процессу оплодотворения у голосемянных и высших споровых до сих пор остаются гипотетичными. Навашин же обнаружил распространенность у ряда растений (березы, лещины, ольхи и некоторых др.) проникания пыльцевой трубочки к зародышевому мешку не через семявход, а через халазу, непосредственно по ткани (халазогамия в отличие от порогамии).
В результате оплодотворения семяпочка превращается в семя с слагающими его частями (см. зародыш растения).
В подготовительных процессах клеточных делений при образовании яйцевых клеток и пыльцы во время образования четверок происходит процесс редукционного деления. Клетки пыльцы возникают путем образования четверок из материнских клеток археспория — группы клеток, закладывающихся на ранних стадиях развития пыльников. У цветковых, таким образом, х-хромосомная генерация, в отличие от споровых, имеет крайне укороченное существование, совершенно утрачивает самостоятельность и остается внутри 2 х-хромосомной генерации.
С точки зрения эволюции растительного царства, картина оплодотворения цветковых растений, взятая в целом, представляет собой глубоко внутреннее оплодотворение в отличие от внешнего оплодотворения у водорослей и высших споровых. Ход эволюции привел и в животном, и в растительном мире к поразительно сходным результатам в связи с приспособлением жизни на суше. И тут и там яйцевая нитка прячется глубоко в материнском организме, будучи защищена таким образом от неблагоприятных условий. Вместе с тем для оплодотворения ее требуется предварительное введение мужских клеток внутрь женского аппарата, что достигается актом совокупления. У самцов животных с этой целью вырабатывается особый копуляционный орган, который и вводится в половые пути самки. Осуществление совокупления достигается сложным механизмом половых рефлексов и инстинктов, в основании которых лежит высокоразвитая у животных нервно-двигательная способность. Вместе с тем у животных окончательное сближение мужских клеток с яйцевой осуществляется путем самостоятельной подвижности первых. У цветковых растений, в связи с общим недостатком у них подвижности, задача совокупления разрешается иначе. Клетки пыльцы, микроспоры, дающие мужской заросток, представляют собой по существу карликовых «самчиков». Образование пыльцевой трубочки при этом соответствует копуляционному органу. «Самчики», за отсутствием собственной подвижности, пассивно переносятся на женский аппарат ветром или насекомыми, в результате целой совокупности приспособлений. Вместе с тем и оплодотворяющие клетки, или ядра, совершенно освобождаются от необходимости самостоятельных движений благодаря тому, что копуляционный орган — пыльцевая трубочка доводит их почти до непосредственного соприкосновения с яйцевой клеткой.
Образование упрощенного мужского заростка в пыльце и женского в семяпочке совершенно еще отчетливо выражено у голосемянных (см. XLV, ч. 3, 177).
Цветов представляет собой не что иное как листостебельный побег, претерпевший глубокий метаморфоз (Гете; см. XIV, 450/51).
Рис. 39. Цветок белой водяной лилии с переходами от лепестков к тычинкам.
Это ясно вытекает из разнообразных фактов. В некоторых нормальных цветах (например, у белой водяной лилии, Nymphaea alba, рис. 39) можно видеть все переходы от наружных, окрашенных в зеленый цвет листочков до тычинок. Еще больше бросаются в глаза подобные переходы и обратные превращения в уродливых цветах: махровых и др. У мака, например, встречается уродливость, при которой тычинки частично превращены в пестики (рис. 40), несмотря на все внешнее кажущееся различие между ними. Нередко встречается полное позеленение всего цветка, когда все части его более или менее превращаются в зеленые листочки. Превращение побега в цветок происходит, вероятно, под влиянием каких-то химических веществ, которые накапливаются в растении к моменту наступления размножения. Опыты Клебса показали, что, изменяя условия питания, удается совершенно изменить ход развития и заставить, например, растение, собиравшееся уже цвести и перешедшее уже к образованью соответственно метаморфизированных побегов, вновь возвратиться к обыкновенному вегетативному росту на неопределенное время. Вообще опыты Фехтинга и др. показывают большую роль внешних условий в формообразовательных процессах цветка. Так, в некоторых случаях зигоморфные цветы удается сделать правильными, устранивши влияние силы тяжести при помощи вращения на клиностате.
Рис. 40. Уродливость мака с тычинками, превращенными в пестики.
Образованно лепестков и их окраска часто в сильнейшей степени зависит от света. С другой стороны, существует обратное взаимоотношение между образованием цветов и вегетативным размножением. Нередко (например, у ряда луковичных) образование цветов в различной степени совершенно вытесняется образованием луковичек и т. п.
Зачатки будущих растений у споровых растений — споры легко разносятся ветром уже в силу своего микроскопического размера. Семена и плоды (см. плод) цветковых растений, вследствие своей тяжеловесности, нуждаются в специальных приспособлениях для своего расселения. Для последней цели цветковые растения пользуются частью опять-таки ветром и насекомыми, частью вырабатывают собственные разбрасывающие механизмы. При расселении ветром семена и плоды бывают снабжены волосистыми хохолками, крыловидными придатками, или просто сильно сплющены, благодаря чему повышается их парусность (отношение поверхности к наибольшему сечению). При расселении животными в одних случаях семена и плоды поедаются животными, благодаря присутствию сочной мякоти, причем самые семена, вследствие твердых оболочек, проходят неизмененными через кишечник и выбрасываются с экскрементами; в других случаях они снабжены разнообразными прицепками, в третьих пристают к ногам, шерсти животных с грязью. Самостоятельное разбрасывание происходит также различными способами. В одних случаях семена просто выбрасываются по инерции из своих вместилищ при встряхивании стебля (баллистический способ), в других — благодаря ссыханию околоплодника развивается значительная механическая сила, ведущая к выбрасыванию семян, в третьих — разбрасывание является результатом тургора сочных тканей плода.
О географическом распространении растений и геоботанике, ископаемых растениях, болезнях растений и растительных сообществах см. фитогеография, фитопалеонтология, фитопатология и фитосоциология.
Литература: К. А. Тимирязев, «Жизнь растения»; В. И. Талиев, «Строение и жизнь растения», 1923; И. П. Бородин, «Краткий учебник ботаники», 1923; В. И. Талиев, «Основы ботаники в эволюционном изложении», 1931; А. Кернер ф. Марилаун, «Жизнь растений», 1890 (перевод с немецкого); И. П. Бородин, «Анатомия растений», 1912; Максимов, «Краткий курс физиологии растений», 1927; Ивановский, «Физиология растений»; В. Л. Комаров, «Практический курс ботаники», 1923; Е. Варминг, «Систематика растений», 1897; «Lehrbuch der Boianik für Hochschulen» begründet v. Ed. Strasburger, 1923; G. Haberlandt, «Physiologische Pflanzenanatomie», 1924; K. Goebel, «Organographie der Pflanzen», 1928; «Handbuch d. Pflanzenanatomie» hrsg. v. K. Linsbauer, W. Вenecke u. L. Jost, «Pflanzenphysiologie», 1923-1924; 0. Kirchner, Е. Loew u. G. Schroeter, «Lebensgeschichte der Blütenpflanzen Mitteleuropas», 1904 и сл.; R. Chodat, «Principes de botanique», 1909; F. Péchoutre, «Biologie florale», 1909; W. Brown, «А textbook of general botany», 1925; Ch. Chamberlain, «Methods in plant histology», 1924.
В. Талиев.
| Номер тома | 36 (часть 1) |
| Номер (-а) страницы | 1 |