Рыбы
Рыбы, сборная группа низших черепных позвоночных (Craniata), которые приспособлены для жизни в воде и отличаются следующими признаками: дыхание жаберное в течение всей жизни; есть только внутреннее ухо; парные конечности, если они есть, построены по типу плавников; непарные плавники поддерживаются скелетными элементами. К рыбам относятся следующие, ныне живущие группы (более подробно о системе рыб см. ниже): круглоротые (Marsipobranchii: миксины, миноги), акулообразные (Selachii), химерообразные (Holocephali), двоякодышащие (Dipnoi) и высшие рыбы (Teleostomi); последние разделяются на Crossopterygii (кистеперые, открытая в 1938 г. Latimeria) и Actinopterygii (лучеперые). Лучеперых условно делят на: Chondrostei (Polypterus, осетры и др.), Holostei (Amia и Lepidosteus) и Теіеоstei (лосось, плотва, сом, окунь и др.). Круглоротые так значительно отличаются от рыб, что их в настоящее время не причисляют к рыбам, называя рыбообразными. В нижеследующем мы, однако, условно описываем их вместе с рыбами.
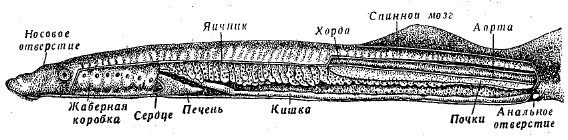
Рис. 1. Внутренности речной миноги (Lampetra fluviatilis).
I. Морфология и анатомия рыб. Внешняя форма. Идеальная форма тела у рыб — веретенообразная, представляющая наиболее удобств для плавания; таковы лосось, сельдь, скумбрия, некоторые акулы. Но есть множество исключений, обусловленных приспособлением к специальному образу жизни: у рыб, живущих на дне, нередко тело уплощенное (камбалы, скаты); миксины, паразиты рыбы, имеют тело червеобразное; угри, морские иглы, Calamoichthys, акула Chlamydoselachus и некоторые другие тоже вытянуты в длину; другие, напротив, укорочены, обладая коротким, но высоким телом, например, солнце-рыба (Моlа) или Psettus. Большим видоизменениям подлежит голова: она то вытянута в пилообразный придаток, как у пилы-рыбы (Pristis) из скатов, то молотообразная, как у рыбы-молота (Sphyrna) из акул, то с лопатообразным рылом, как у лопатоноса (Scaphirhynchus) из осетровых. Рот у Selachii почти всегда нижний, у Chondrostei всегда нижний, у Teleostei обычно конечный. Ротовое отверстие иногда очень велико (у акул, у некоторых глубоководных Teleostei), у иных — очень мало (Mormyridae). Заднепроходное отверстие лежит обычно на границе туловища и хвоста, перед заднепроходным плавником, но иногда бывает перенесено далеко вперед, на горло (у американского семейства Gymnotidae). Глаза на боках головы, но иногда наверху (Uranoscopus); у камбал оба глаза смещены на одну сторону, правую или левую. У некоторых глубоководных и пещерных рыб глаза зачаточны, также у миксины. Носовые отверстия у акулообразных и химер лежат на нижней стороне головы; у Dipnoi каждый обонятельный мешок снабжен двумя отверстиями: одно наружное, лежит на нижней стороне рыла, другое, внутреннее, ведет в полость рта. У круглоротых одно непарное носовое отверстие. У прочих рыб органы обоняния на верхней стороне головы, обычно в виде парных отверстий с каждой стороны. Голова заканчивается жаберной областью. У акул 5—7 жаберных отверстий на боках тела, у скатов 5 внизу, у химер есть складка кожи в виде жаберной крышки (но без специального скелета). У высших рыб и Dipnoi есть только одно наружное жаберное отверстие с каждой стороны, прикрытое жаберной крышкой, состоящей из костей. Из круглоротых у миног 7 пар жаберных отверстий (рис. 1), у миксины — одна пара, отнесенная далеко назад (у других Мухіnі 5—15 пар жаберных отверстий). Почти у всех акул, у скатов, у некоторых осетровых и др. есть позади глаз отверстие, называемое брызгальцем (spiraculum); это зачаточная жаберная щель между челюстной и подъязычной дугами. На боках тела у большинства рыбы мы находим ряд чешуй, пронизанных отверстиями; это т. н. боковая линия, своеобразный орган чувств, служащий для ориентировки в воде. Плавники состоят из лучей, которые могут быть членистыми и нечленистыми; членистые лучи бывают ветвистыми и неветвистыми; нечленистые могут иметь вид колючек. Плавники делятся на 1) непарные, или вертикальные: спинной, хвостовой, анальный (заднепроходный); спинных и анальных плавников иногда бывает несколько (например, у трески — 3 спинных и 2 анальных); 2) парные: грудные и брюшные. У круглоротых совершенно нет парных плавников. У некоторых низших Teleostei (например, у лосося, сига, многих сомов) за спинным плавником бывает небольшой плавник обычно без лучей (жировой плавник). Грудные плавники лежат за жаберными отверстиями; редко вторично отсутствуют (Muraenidae, некоторые Stomiatoidei, см. рис. 19); у скатов достигают громадного развития. Брюшные плавники лежат или за грудными на брюхе (сельдь, плотва, щука), или под грудными (окунь), или впереди их (треска), иногда даже на подбородке (Ophidium), иногда срастаются вместе, образуя присоску (бычки: Gobius), нередко отсутствуют (у угрей, морских игл). У самцов всех Selachii и Holocephali задние лучи брюшных плавников превращены в парный совокупительный орган (птеригоподии). Хвостовой плавник у ныне живущих рыб бывает: 1) первично-симметричный (протоцеркальный), когда он располагается по оси тела: круглоротые; 2) несимметричный (гетероцеркальный), когда конечная часть хвоста загибается вверх: акулы (см. ниже; рис. 15), осетровые; 3) ложно-симметричный (гомоцеркальный), т. е. наружно симметричный, но анатомически асимметричный: большинство Teleostei (рис. 2 и 3); 4) вторично-симметричный (дифицеркальный): Dipnoi, Fierasfer. Очень редко хвостовой плавник отсутствует (например, у морского конька).
Рис. 2. Скелет гомоцеркального хвостового, плавника сазана (Cyprinus саrріо). К последнему, неполному позвонку прикрепляются hypuralia; а — уростиль.
Величина рыб сильно варьирует: среди них есть и самые мелкие, и самые крупные из современных позвоночных. У Pandaka pygmaea, бычка (Gobiidae) с Филиппинских островов, зрелые самцы имеют в длину от 7,5 до 9 мм, некоторые Cyprinodontiformes достигают всего 15—20 мм. Напротив, некоторые акулы достигают 15 м (Rhinodon typicus, Cetorhinus maximus). Обычно самцы рыб мельче самок; это зависит от того, что самцы обычно растут медленнее и раньше достигают половой зрелости. Половой диморфизм нередко выражен у рыб весьма резко; у многих самцы и самки окрашены различно (Labrus). В брачном наряде самцы иногда сильно отличаются от самок: у самцов многих карповых на теле появляются эпителиальные бугорки, у самцов горбуши (Oncorhynchus gorbuscha) на спине развивается горб, во рту вырастают сильные зубы (в меньшей мере эти признаки развиваются и у самок горбуши). У лосося (Salmo salar) есть самцы двух родов — крупные (а лосось достигает веса 30 кг и выше) и мелкие; последние могут иметь зрелые половые продукты при длине всего в 10 см; самки же лосося никогда не бывают половозрелыми такой длины. Что касается влияния внешних условий на величину рыб, то можно указать, что морские рыбы крупнее солоновато-водных; так, балтийская треска мельче океанической. Живущие в более или менее замкнутых бассейнах мельче живущих в больших: фьордовая треска мельче океанической (мигрирующей). Рыбы северной части Тихого океана крупнее, чем рыбы тех же родов, живущие в северной части Атлантического океана. Многие северные рыбы крупнее южных; так, среди 35 тыс. камбал Platessa platessa, исследованных в датских проливах, только одна была величиной в 49 см, тогда как на севере Норвегии встречаются особи той же камбалы в 70 и даже 80 см длиной.
Рис. 3. Хвостовая часть позвоночника форели (Salmo trutta fario): d — верхние остистые отростки; е — расширенные верхние остистые отростки; f — нижние остистые отростки; g - расширенные нижние остистые отростки; h — hypuralia; i — уростиль.
Кожный скелет. Тело рыб бывает или голым (круглоротые, многие сомы и др.) или, чаще, покрыто чешуей, шипиками или пластинками. Различают чешуи нормального типа, ганоидные и плакоидные. 1) Чешуи нормального типа представляют собой тонкие костные пластинки, у высших Teleostei лишенные костных клеток; они бывают циклоидными, т. е. с гладким задним (свободным) краем, как у сазана, или ктеноидные — с зазубренным задним краем, как у окуня. Как правило, циклоидные чешуи встречаются у более низко организованных Teleostei — у имеющих абдоминальные брюшные плавники (сельдь, лосось, плотва, лещ), тогда как у более высоко организованных рыбы, например, у судака, чешуи ктеноидные. Однако бывает, что у одной и той же рыбы на разных частях тела имеются чешуи обоих типов, например у некоторых бычков (Gobiidae); у камбал из рода Liopsetta — у самок циклоидные чешуи, у самцов ктеноидные. 2) Ганоидные чешуи представляют собой толстые, плотные пластинки, ромбической или квадратной формы, расположенные косыми рядами. Эта форма чешуй, весьма обычная у ископаемых рыб, из ныне живущих свойственна лишь Lepidosteus и Polypterus; у осетровых подобные (но только по форме) чешуи имеются лишь на верхней лопасти хвостового плавника. Сверху ганоидная чешуя покрыта толстым слоем плотного бесклеточного эмалеподобного вещества, называемого ганоином. Под ганоином у Lepidosteus лежат слои костного вещества (изопедин). Ганоин, в отличие от эмали, не эпителиального происхождения, а соединительнотканного. Следует еще прибавить, что у Polypterus и Lepidosteus на чешуях встречаются зубчики, построенные совершенно по типу кожных зубов (см. ниже). У Polypterus между ганоином и изопедином залегает слой с сосудистыми каналами и дентинными трубочками (космин). 3) Плакоидные чешуи (у Selachii) — не что иное, как кожные зубы; они состоят из дентина (соединительнотканного происхождения) и эмали (из эпидермиса). У многих Teleostei на коже бывают пластинки (например, у панцирных сомов); это — кожные кости.
Органы свечения, встречающиеся у некоторых морских рыб, есть, по-видимому, видоизмененные кожные слизеотделительные железы.
Окраска рыб зависит: 1) от, особых пигментных клеток, содержащих красное, оранжевое, желтое и черное начало и называемых хроматофорами; 2) от клеток, заключающих кристаллы гуанина и называемых иридоцитами; отражаясь от иридоцитов и подвергаясь интерференции, свет дает начало массе цветовых оттенков. Хроматофоры и иридоциты расположены как на чешуях, так и под ними. Многие рыбы меняют окраску в зависимости от окраски окружающей среды (тюрбо, Rhombus maximus), от возраста и состояния половой зрелости (лососи).
Скелет. У круглоротых скелет всю жизнь остается хрящевым или даже перепончатым. У Selachii и Holocephali хрящ пропитывается известью, настоящих же костей у них нет; у Dipnoi и Teleostomi есть в большем или меньшем количестве кость. У круглоротых, Holocephali, Dipnoi и Chondrostei хорда остается в течение всей жизни («вязига» осетровых), у остальных — более или менее полно заменена телами позвонков. Позвонки у Teleostei двояковогнутые (амфицельные) и лишь у одного Lepidosteus спереди выпуклы, сзади вогнуты (опистоцельные). Ребра у Polypterus и у большинства Teleostei имеются и нижние, и верхние (верхние ребра у Teleostei обычно носят название epipleuralia); у круглоротых, скатов, химер и морских игл (Syngnathidae), у Lophiiformes и некоторых других Teleostei совсем нет ребер. У прочих рыб ребра нижние.
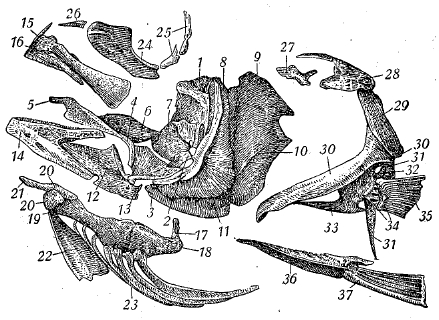
Рис. 3-А. Morone saxatilis. Семейство Serranidae. Кости боков черепа, плечевого пояса, пояса грудных и брюшных плавников.
1. Hyomandibulare. 2. Symplecticum. 3. Quadratum. 4. Pterygoideum (ectopterygoideum). 5. Palatinum. 6. Mesopterygoideum (entopterygoideum). 7. Metapterygoideum. 8. Praeoperculum. 9. Operculum. 10.Suboperculum. 11. Interoperculum. 12. Articuiare. 13. Angulare. 14. Dentale. 15. Maxillare. 16. Praemaxillare. 17. Stylohyale (interhyale). 18. Epihyale. 19. Ceratohyale. 20. Hypohyale (два с каждой стороны). 21. Glossohyale (basihyale). 22. Urohyale. 23. Radii branchiostegi. 24. Praeorbitale. 25. Infraorbitalia («suborbitalia»). 26. Nasale. 27. Supratemporale (окостенение вокруг канала боковой линии). 28. Posttemporale. 29. Supracleithrum. 30. Cleithrum. 31. Postcleithrum. 32. Scapula. 33. Coracoideum. 34. Radialia грудного плавника. 35. Плавниковые лучи грудного плавника. 36. Тазовые кости. 37. Плавниковые лучи брюшного плавника.
Скелет головы. Опишем сначала череп окуня (см. рис. 3-А), а) Черепная коробка. В затылочной области мы находим basioccipitale, 2 occipitale laterale (цифра 2 означает, что кость парная), supraoccipitale, 2 opisthoticum (intercalate); в области уха: 2 prooticum, 2 pteroticum, 2 epioticum, 2 sphenoticum (postfrontale); сверху черепа: 2 parietale, 2 frontale, 2 ethmoidale laterale (praefrontale), mesethmoideum, 2 nasale. Внизу черепа спереди сошник (vomer), сзади parasphenoideum; к последнему примыкает basisphenoideum. Есть 2 alisphenoideum. Под глазом несколько подглазничных (infraorbitalia); из них передняя, самая крупная называется рrаеorbitale (2). б) Верхняя и нижняя челюсть с подвеском. Верхняя челюсть состоит из 2 praemaxillare (покрытых зубами) и 2 maxillare; нижняя челюсть — из 2 dentale (с зубами), 2 articulare и 2 angulare. На внутренней стороне articulare лежит хрящевой тяж — остаток мекелева хряща. С верхней челюстью соединен ряд небокрыловидных костей: 2 palatinum (с зубами), 2 ectopterygoideum, 2 metapterygoideum, 2 entopterygoideum. С нижней челюстью сочленяется 2 quadratum, которая при помощи 2 symplecticum и 2 hyomandibulare служит для прикрепления нижней челюсти к черепу. На hyomandibulare лежит снаружи praeoperculum. Каждая жаберная крышка состоит из 3 костей: operculum, interoperculum, suboperculum. в) Подъязычная дуга, начинаясь hyomandibulare, состоит с каждой стороны из stylohyale (или interhyale) и подъязычной (epihyale + ceratohyale + 2 hypohyale). На epihyale и ceratohyale сидят всего 7 лучей жаберной перепонки (radii branchiostegi) с каждой стороны. В язык вдается непарная язычная (glossohyale, или basihyale, или linguale); от места соединения правой и левой подъязычных дуг отходит назад непарная urohyale. г) Жаберных дуг 5 с каждой стороны. В плане они построены, как и подъязычная: непарный элемент представлен basibranchiale, или copula, справа и слева от него: hypobranchiale, ceratobranchiale, epibranchiale и pharyngobranchiale. Но задние дуги имеют упрощенное строение, а последняя, 5-ая, состоит с каждой стороны из одной так называемой нижнеглоточной кости (os pharyngeum inferius), покрытой зубами; на этой дуге нет жаберных лепестков.
У круглоротых нет элементов, соответствующих верхней и нижней челюстям; у миноги рот окаймлен большим кольцевидным хрящом. У акул череп представляет большую хрящевую коробку; верхняя челюсть состоит из пары небоквадратных хрящей (palatoquadratum), усаженных зубами; нижняя, тоже покрытая зубами, представлена парой хрящей (мекелевы хрящи). У Holocephali небоквадратный хрящ плотно прирастает к основанию черепа; такой способ соединения верхней челюсти с черепом, характерный для четвероногих, называется автостилией. Он встречается из рыбы еще только у Dipnoi, но в отличие от Holocephali в черепе двоякодышащих есть окостенения; в области рта у Dipnoi появляются кожные кости; правда, ни maxillare, ни praemaxillare у них нет, но в нижней челюсти у Neoceratodus есть кости (с внутренней стороны нижней челюсти есть большое praearticulare, или spleniale, с сильными зубами — по одному на каждом praearticulare).
Плечевой пояс у окуня (см. рис. 3-А) прикреплен к черепу при помощи posttemporale; в состав пояса входят с каждой стороны supracleithrum, cleithrum, две postcleithrum, scapula (лопатка), coracoideum. К scapula причленены 4 основных косточки (radialia), на которых сидят лучи грудного плавника. Тазовой пояс у окуня состоит из двух костей, назади сросшихся (ossa pubis, или basipterygium); на этих костях, соответствующих лопатке и коракоиду, сидят плавниковые лучи.
У акул плечевой пояс хрящевой, без разделения на отдельные элементы. У Dipnoi, Polypterus и др. Chondrostei есть два кожных окостенения, соответствующих ключице Teleostei: верхняя ключица (cleithrum) и нижняя (clavicula); у Holostei (рис. 4) и Teleostei (рис. 5) остается только верхняя ключица (cleithrum; ее, однако, нередко называют clavicula). У Neoceratodus скелет грудного плавника весьма своеобразен (бисериального типа): к членистой хрящевой оси справа и слева причленяются хрящевые лучи, по большей части сегментированные. У Holocephali и Teleostomi пояс задней конечности представлен парой элементов (рис. 6), у Selachii и Dipnoi — непарным хрящом. К поясу задней конечности у всех, кроме Teleostei, прикреплено много radialia; у Teleostei брюшные radialia отсутствуют или зачаточны.
Электрические органы есть производное мышечной ткани; они есть у некоторых скатов (Torpedo, Raja), у Electrophorus (из Gymnotoidei), у Mormyridae. Только у нильского электрического сома (Malapterurus electricus) электрические органы есть производное эпидермиса.
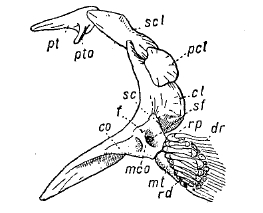
Рис. 4. Плечевой пояс и скелет правого грудного плавника у Аmіа саlѵа изнутри: сl — cleithrum; со — коракоид; dr — плавниковые лучи; f — отверстие для нерва; mco — мезокоракоид; mt — metapterygium; pcl — postcleithrum; pt — posttemporale; pto — отросток posttemporale, идущий к opisthoticum (intercalate); rd и rp — проксимальное radiale; sc — лопатка; scl — supracleithrum; sf — лопаточное отверстие. Хрящ обозначен точками.
Нервная система. У миксин ни промежуточный мозг, ни мозжечок не обособлены, но есть полушария большого мозга; у миног есть промежуточный мозг, снабженный двумя непарными теменными глазами; мозжечок слабо развит. У Selachii и Holocephali мозжечок сильно развит; париетального глаза нет. У Teleostei (рис. 7) бросается в глаза преобладание среднего мозга, передний мозг едва развит и имеет эпителиальную крышу; мозжечок хорошо развит. У Dipnoi полушария большого мозга сильно развиты, но мозжечок мал. Головных нервов у настоящих рыбы 10 пар. Из органов чувств упомянем об органах боковой линии, свойственных всем рыбам и водным амфибиям. Орган обоняния у круглоротых вступает в связь с нижним мозговым придатком (hypohpysis). У рыб есть только внутреннее ухо. У миксин и миног два полукружных канала, у настоящих рыб – три.
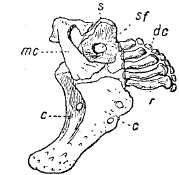
Рис. 5. Плечевой пояс и скелет правого грудного плавника лосося (Salmo salar) изнутри: с – коракоид; dc – дистальные хрящи скелета грудного плавника; mc – мезокоракоид; r – radiale; s – лопатка; sf – лопаточное отверстие.
Кишечник. Зубы у круглоротых роговые, у Selachii и Teleostomi построены по типу плакоидных чешуй. Некоторые, например, карповые (Cyprinidae), не имеют во рту зубов, но зато у них есть зубы на нижнеглоточных костях. У осетровых (Асіреnseridae) совсем нет зубов (но у мальков есть). «Язык» у круглоротых снабжен мощными мышцами. Мышцы в языке есть у Polypteridae и Dipnoi, у прочих мышц в языке совсем нет. Иногда (Muraenidae) языка не бывает. Обычно у рыбы можно различить пищевод, желудок и кишку.
При начале кишки в нее у многих рыб впадают слепые отростки, т. н. пилорические придатки (рис. 8). У миног (Petromyzones) и всех настоящих рыб, кроме Teleostei, есть в кишке спиральный клапан — складка всех или нескольких слоев кишечной стенки. У Selachii и Dipnoi есть клоака (у Teleostei она бывает как исключение, например, у самки морской иглы, Nerophis aequoreus). Глоточная часть кишечника у всех рыбы пронизана жаберными щелями. Свежая вода, омыв жаберные лепестки и окислив венозную кровь, принесенную к жабрам, выходит через жаберные щели наружу. В промежутках между жаберными щелями находятся хрящевые или костные жаберные дуги (у круглоротых жаберный скелет залегает поверхностно). У Теіеоstomi 5 внутренних жаберных щелей или менее; снаружи они покрыты жаберной крышкой, поддерживаемой костями.
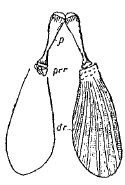
Рис. 6. Тазовой пояс и брюшные плавники у Amia calva: dr – плавниковые лучи, р – тазовая кость, prr – один из radialia.
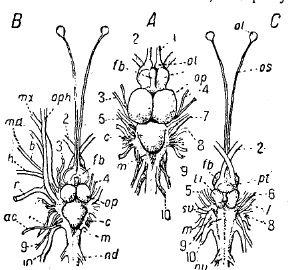
Рис. 7. А — мозг щуки (Esox Iucius) со спинной стороны, В — мозг трески (Gadus morhua) со спинной стороны, С — мозг трески с брюшной стороны: 1 — n. olfactorius, 2 — n. opticus, 3 — n. oculomotorius, 4 — n. trochlearis, 5 — n. trigeminus, 6 — n. abducens, 7 — n. facialis, 8 — n. acusticus, 9 — n. glossopharyngeus, 10 — n. vagus, ac — r. accessorius, b — r. buccalis, h — r. hyomandibularis, md — r. mandibularis, mx — r. maxillaris, oph — r. ophthalmicus superior, r—r. dorsalis recurrens, c — мозжечок, fb — передний мозг (в A epiphysis), li — lobus inferior, m — продолговатый мозг, ol — обонятельная лопасть, op — lobus opticus, os — tractus olfactorius, pt — нижняя мозговая железа.
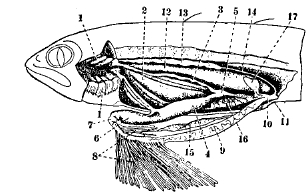
Рис. 8. Сельдь Alosa fallax, вскрытая с левой стороны: 1 — перерезанные жаберные дуги, 2—вскрытый желудок, 3 — верхняя стенка желудка, 4 — нижняя стенка желудка, 5 — задняя связка, поддерживающая желудок, 6 — пилорическая часть желудка, 7 — пилорический клапан, 8 — пилорические придатки, 9 — кишка, 10 — конечный отдел кишки, 11 — заднепроходное отверстие, 12 — плавательный пузырь, 13 — почка, 14 — мочеточник, 15 — селезенка и поджелудочная железа, 16 — правый семенник, 17 — часть отрезанного левого семенника.
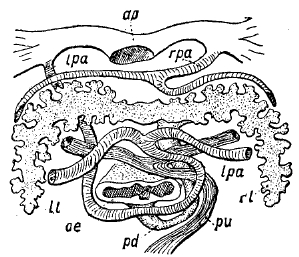
Рис. 9. Разрез через Protopterus (схема): ао — место соединения дуг аорты в спинную аорту, ll — левая лопасть плавательного пузыря, Іра — левая легочная артерия, ое — пищевод, pd — ductus pneumaticus, pa — легочная вена, rl— правая лопасть плавательного пузыря, rра — правая легочная артерия.
У Holocephali жаберная область прикрыта складкой кожи, поддерживаемой хрящевыми лучами подъязычной дуги. У круглоротых жабры имеют вид мешков, сообщающихся как с наружной средой, так и с глоткой (у взрослых миног мешки открываются не в глотку, а в особый непарный жаберный канал, лежащий под пищеводом). У Selachii обычно 5 жаберных отверстий (у Heptanchus — 7). Жаберные перегородки у Selachii хорошо развиты, у костистых — зачаточны. 5-ая жаберная дуга у Teleostomi лишена жабр (как и у акул, имеющих 5 жаберных отверстий); таким образом, у них не более 4 пар жабр, но может быть и меньше (у Amphipnous из Symbranchiformes 1 пара). На передней стороне жаберных дуг имеются хрящевые или костные тычинки или бугорки, т. н. жаберные тычинки, имеющие целью препятствовать ускользанию пищи вместе с водой наружу, через жаберные щели, и защищать жабры от поранения. У некоторых рыбы (например, у Аnаbas) в жаберной полости развиваются придаточные органы дыхания. У других рыб вспомогательным органом дыхания служит кожа (угорь), или кишечник (вьюн), или плавательный пузырь (см. ниже).
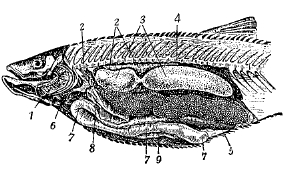
Рис. 10. Голавль (Leuciscus cephalus), вскрытый с левой стороны: 1 — жабры, 2 — почки, 3 — плавательный пузырь, 4 — позвоночник, 5 — яичник, 6 — сердце, 7 — кишечник, 8 и 9 — печень.
У Dipnoi (рис. 9) и Teleostomi (рис. 10) есть плавательный пузырь в виде полого мешка, лежащего обычно над кишечником. У молодых он всегда, через ductus pneumaticus, соединен с пищеводом, а у многих взрослых Teleostei (окунь и мн. др., т. н. закрытопузырные) соединение с кишечником прекращается. У многих придонных рыб плавательный пузырь исчезает (взрослые камбалы, многие Cottidae и др.). У Polypteridae и Dipnoi плавательный пузырь открывается с брюшной стороны, т. е. там же, где и легкие четвероногих. У Polypterus плавательный пузырь парный, сильно ячеистый, похож на легкие. Ячеистым бывает он и у Holostei и некоторых Teleostei (Erythrinus и др.). Роль плавательного пузыря — поддерживать в теле рыбы давление, равное удельному весу воды; у иных он, как мы видели, делается органом дыхания, у некоторых служит для произведения звуков (Sciaena, сомы Doras, Auchenipterus).
Кровеносная система. Сердце у рыб лежит сейчас же за последней парой жабр, занимая гораздо более переднее положение, чем у четвероногих. Сердце рыбы (рис. 11) состоит из желудочка и предсердия, лежащего на спинной стороне желудочка. Предсердие получает венозную кровь и посылает ее через посредство желудочка в жабры, откуда она, уже окисленная, разносится по всему телу. Впереди желудочка у некоторых рыб (Selachii, Holocephali, Dipnoi, Chondrostei, Holostei, низшие Teleostei) имеется мускулистый отдел, т. н. артериальный конус, снабженный клапанами. От артериального конуса, а если его нет, то прямо от желудочка отходит непарный артериальный ствол (truncus arteriosus), или брюшная аорта, идущая под жабрами по средней линии тела. От брюшной аорты направо и налево отходят к каждой жабре жаберные артерии, которые в жаберных лепестках распадаются на капилляры; здесь венозная кровь окисляется и попадает в жаберные вены, которые под позвоночником впадают в непарную спинную аорту. Венозная кровь попадает в сердце из головы через парные яремные вены (ѵ. jugulares, или cardinales anter.), из туловища — через парные кардинальные вены; те и другие в области сердца соединяются и образуют с каждой стороны по верхней полой вене (vena cava anterior, иначе ductus Cuvieri), которые впадают в венозную пазуху (sinus venosus), открывающуюся в предсердие. В венозную же пазуху впадают и печеночные вены. У Dipnoi, обладающих и легочным, и жаберным дыханием, намечается разделение предсердия на две половины: правую, получающую венозную кровь, и левую — артериальную. У некоторых из Teleostei, способных к воздушному дыханию, тоже намечается смешение в сердце венозной и артериальной крови (например, у Monopterus).
Мочевая система (ср. XXIX, приложение 3894/914). У рыб во взрослом состоянии функционируют у одних первичные почки (pronephros), у других — и сюда относится подавляющее большинство рыб — вторичные (mesonephros). Первичная почка представляет собой ряд посегментно расположенных канальцев, открывающихся с одной стороны в полость тела при посредстве воронок, а с другой — в общий выводной проток (пронефрический проток); правый и левый выводной протоки впадают в клоаку. Перед воронками лежат клубочки из капилляров (glomeruli). Выделяемая клубочками жидкость поступает непосредственно в полость тела, откуда она, при посредстве ресничек воронки, переходит в почку. Вторичная почка развивается кзади от первичной, но в качестве выводного протока пользуется пронефрическим протоком. У Selachii пронефрический проток расщепляется по своей длине на два протока: один из них (вольфов проток) служит выводным каналом для почек, а другой (мюллеров проток) у самок превращается в яйцевод, выносящий яйца наружу. Первичные почки имеются у всех личинок круглоротых и у мальков рыбы, но во взрослом состоянии они функционируют, и то лишь отчасти, у некоторых круглоротых (Bdellostomatidae), а затем у Dipnoi, осетровых, Polypterus, Holostei, у некоторых Teleostei, каковы: Zoarces, Fierasfer, Cottus gobio, Dactylopterus, Lepadogaster и др. У взрослых круглоротых, за исключением Bdellostomatidae, имеются вторичные почки, а мочеточники представляют собой пронефрические протоки; сзади мочеточники впадают в мочевой синус, который открывается наружу позади анального отверстия.
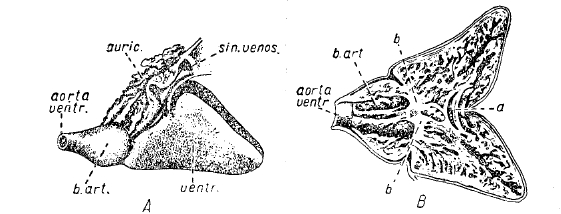
Рис. 11. Сердце кумжи (Salmo trutta): А — невскрытое, В — вскрытое: а — клапан на месте перехода предсердия в желудочек, aorta ventr. — брюшная аорта, auric. — предсердие, b — клапаны, b. art. — bulbus arteriosus, sin. venos — венозный синус, ventr. — желудочек.
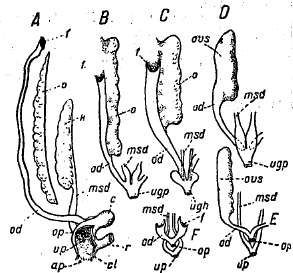
Рис. 12. Схемы женских мочеполовых органов: А — Protopterus, В — Polypterus, С — Аmіа, D — Lepidosteus, Е — типичная костистая, F — Salmonidae: ар — абдоминальная пора, с — клоакальный пузырь, сl — клоака, f — воронка яйцевода, открывающаяся в полость тела, k — почка (mesonephros), msd — мочеточник, о — яичник, od — яйцевод, ор — половой сосочек или половая пора, ovs — замкнутый яичник, r — прямая кишка, ugp — мочеполовой сосочек, up — мочевое отверстие.
Половые органы. У круглоротых половые железы обычно непарны, выводных протоков нет, и половые продукты попадают прямо в полость тела, откуда выносятся наружу через брюшную пору, называемую половой. Миноги раздельнополы, у миксин же задняя часть половой железы функционирует как семенник, передняя — как яичник. У настоящих рыб яичники или лежат свободно в полости тела, или представляют собой мешки, окруженные оболочкой (рис. 12). Первый тип свойственен Selachii, Dipnoi, Polypterus, осетровым, некоторым Teleostei, например, Salmonidae, Osteoglossidae, угрям, Misgurnus (рис. 13). Замкнутые яичники мы находим у большинства Teleostei, а также у Lepidosteus. У Selachii, у осетровых, Аmіа и Lepidosteus выводным протоком для семенников служат выводные протоки почек (вольфовы протоки); между тем Polypterus, Protopterus и Teleostei имеют обособленные семяводы, не стоящие в связи с выводными протоками почек (рис. 14). У Ceratodus и Lepidosiren отношения семяводов к почкам напоминают то, что наблюдается у осетровых.
Рис. 13. Вьюн, Misgurnus fossilis, вскрытый слева: 1 — жабры, 2 — плечевой пояс, 3 — плавательный пузырь, 4 — печень, 5 — яичник, 6 — почка, 7 — язык, 8 — грудной плавник, 9 — сердце, 10 — селезенка, 11 — кишка.
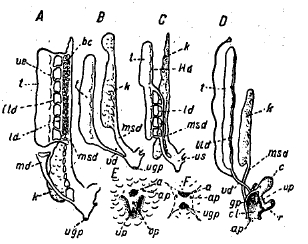
Рис. 14. Схемы мочеполовых органов у самцов: А — Acipenser (таково же строение у Lepidosteus и Аmіа, но у них нет мюллерова протока, md), В — Teleostei, С — Polypterus, D — Protopterus, Е — мочеполовой сосочек самки Salmo, F — то же у самца Polypterus: а — анальное отверстие, ар — абдоминальная пора, bс — почка, с — клоакальный пузырь, gp — половой сосочек, k — почка (mesonephros), Id — продольный, выводной проток, ltd — продольный выводной проток семенника; md — мюллеров проток, msd — мочеточник, ор — половая пора, r — прямая кишка, t — семенник, ugp — мочеполовая пора, up — мочевая пора, us — мочеполовой синус, vd — vas deferens, ve — vas efferens.
II. Образ жизни. Размножение. У многих рыб (может быть у всех) число самцов равно числу самок (у горбуши, сельди, осетровых, камбал). Очень крупные яйца — у акул и Holocephali (у Callorhynchus 25 см), у Teleostei обычно не свыше 5—6 мм, но у сома Arius — 17—18 мм. Количество икры различно: у Моlа длиной в 1 ½ м 300 миллионов икринок, у крупной трески свыше 9 миллионов, у сельди — 29—127 тысяч, у байкальского Asprocottus herzensteini, длиной 60 мм, всего 19 штук. Как правило, чем крупнее особь, тем икры у нее больше. У воблы:
3 лет – 17,1 тыс. икринок;
4 – 35,3;
5 – 50,0;
6 – 88,2;
7 – 136,5.
Одни рыбы мечут икру в теплое время года (карповые, осетровые), другие — в холодное (лосось, сиги, налим). Самцы вообще созревают раньше самок: в Северном море у камбалы (Platessa platessa) самцы делаются половозрелыми в конце 3-го года, самки—в конце 5-го; но у некоторых (вобла) — одновременно. Некоторые рыбы после икрометания сплошь погибают (угорь, тихоокеанские лососи, миноги, некоторые каспийские сельди). Оплодотворение обычно происходит в воде, куда самец и самка выпускают половые продукты; но у всех акулообразных и Holocephali, а также у некоторых Teleostei происходит совокупление; у таких рыб развитие яйца совершается в теле матери, причем обычно наблюдается живорождение [бельдюга (см.), Zoarces viviparus]. Время, потребное для вылупления из икры, зависит от температуры: чем она выше, тем развитие идет скорее; так, у горбуши при 8,1° вылупление происходило на 61—84-й день, а при 6,3° — на 87—100-й. Рыбы, мечущие икру летом, развиваются гораздо быстрее: окунь вылупляется на 4-й, 5-й день. Многие рыбы заботятся о потомстве: самец колюшки строит гнездо и охраняет яйца и молодь (см. гнезда, XV, 234/35); самцы морских игл и коньков имеют яйцеприемник на брюхе, куда самка откладывает яйца (о сомах см. XL, 152/53). Самец новогвинейской Kurtus gulliveri (см. ниже, рис. 32) вынашивает икру на затылке, где она прикрепляется при помощи особых нитей к костяному отростку supraoccipitale; самец носит икру вплоть до вылупления мальков. Самец пинагора (Cyclopterus lumpus), обыкновенного на берегах Мурмана, оберегает икру, отложенную на камнях; когда мальки выклюнутся, они первое время прикрепляются к брюху самца при помощи своих брюшных плавников, видоизмененных в присоску. У южно-американского сома Aspredo (cм. сомовые, XL, 152/53) яйца прикрепляются, в виде одного слоя, к брюху самки, а также к нижней поверхности головы и парных плавников; каждое яйцо получает, при помощи сосудов, питание из тела матери. О размножении горчака см. XVI, 91. Тропические азиатские пресноводные рыбы из семейства Anabantidae (Betta, Trichogaster и др., охотно разводимые в аквариумах) устраивают оригинальные гнезда из пены. Они заглатывают на поверхности воды воздух, окружают во рту пузырьки воздуха особым слизистым секретом и затем выпускают их снова наружу, на поверхность воды, где, таким образом, получается как бы гнездо из пены до 10 см в диаметре.
В это гнездо откладываются оплодотворенные яйца. Самец заботится как об икре, так и о мальках, которые вылупляются через 1 ½ — 2 суток. Через 8 дней гнездо начинает растворяться, и мальки покидают его. Помеси среди рыбы — не редкость (стерлядь и осетр, форель и голец, карась и сазан и др.).
Сожительство и паразитизм. Некоторые рыбы сожительствуют с другими рыбами или даже с беспозвоночными. Так, у берегов Новой Гвинеи один великолепно окрашенный вид Amphiprion (из семейства Pomacentridae) живет внутри крупной морской анемоны. Fierasfer (из Ophidioidei) живет внутри голотурий и морских звезд. У нас в Баренцевом море мальков пикши (Меlаnogrammus aeglefinus) находят под колоколами крупных медуз Суаnеа; здесь они живут до поздней осени. Мелкие червеобразно удлиненные южно-американские сомики Vandellia живут в жаберной полости крупных сомов, питаясь кровью последних. Карликовые самцы некоторых рыб-удильщиков (Ceratoidei) паразитируют на своих самках, питаясь их кровью (рис. 36).
Рост. Рост чешуй у костистых рыбы идет так, что ежегодно по периферии можно видеть новый слой вещества, при этом летом прирост идет быстро, а зимой медленно; получаются два кольца, отличающиеся шириной и консистенцией; оба кольца соответствуют одному годичному периоду. Чешуя, а также позвонки, кости жаберной крышки, отолиты — являются удобнейшими объектами для исследования возраста. Оказалось, что сельди у берегов Норвегии живут до 18 лет, камбалы (Platessa platessa) в Северном море — до 30 лет, амурская белуга, или калуга (Huso dauricus) весом в 650 кг — до 50—55 лет. У лосося по чешуе можно узнать, в каком году он метал икру: у половозрелых лососей в реке края чешуи подвергаются эрозии, и след от этого сохраняется и тогда, когда по периферии отложится новое кольцо. По чешуе можно было узнать, что наш северный лосось, или семга (Salmo salar), впервые входит для икрометания в реки из моря обычно на 5-м или 6-м году от роду; выметав икру, он скатывается в море и возвращается снова в реку или в следующем году или через год. Вообще лосось живет, как правило, не более 8—9 лет и за это время мечет икру до 3, редко до 4 раз.
По питанию рыб делят (более или менее условно) на 1) хищных (щука, судак, лосось), 2) мирных, т. е. питающихся планктоном или мелкой береговой фауной (морская сельдь, уклейка, стерлядь), 3) растительноядных (подуст), 4) всеядных, т. е. питающихся мелкими беспозвоночными, а также растениями (сазан).
Отношение к среде. Сообразно с тем, где рыбы мечут икру — в пресной воде или в соленой, — их можно разделить на две группы: пресноводных и морских (солоноватоводных). Вне времени нереста, во время нагула, рыбы могут вести самый различный образ жизни. Одни рыбы (жилые) остаются всю жизнь в той же среде, где они вывелись: пресноводные в пресной, морские в морской, не совершая при этом больших передвижений. Из числа пресноводных рыб сюда принадлежат, например, карась, горчак, линь, красноперка, уклейка, вьюн, щиповка, подкаменщик, налим, щука, окунь и др. Другие жилые рыбы совершают обширные миграции (см.), не покидая, однако, своей обычной среды (т. е. не переходя из морской воды в пресную или из пресной в морскую). Таковы, например, среди морских рыб треска, пикша, скумбрия (макрель), анчоус, сардины, сельдь; среди пресноводных — некоторые озерные сиги, входящие для нереста из озер в реки, также ладожский лосось (Salmo salar morpha sebago), поднимающийся для икрометания в притоки Ладожского озера, наконец, многие речные рыбы, которые с целью нереста совершают более или менее значительные миграции вверх по реке. Но есть большая категория рыб, называемых проходными, которые для икрометания покидают свою среду и переходят из моря в реки или обратно из рек в море. К числу уходящих из рек в море для нереста принадлежит очень небольшое число рыб — речной угорь (Anguilla), речная, камбала (Pleuronectes flesus). Но зато немало имеется таких проходных рыб, которые с целью размножения подымаются из морей в реки; сюда относятся лосось (Salmo salar), кумжа (Salmo trutta), миноги: речная (Lampetra fluviatilis), японская (L. japonica), каспийская (Caspiomyzon wagneri), представители рода Oncorhynchus (кета, горбуша, чавыча и др.), затем белуга, севрюга, русский осетр (Acipenser güldenstädti), сельдь-черноспинка (Caspialosa kessleri), волжская сельдь (Caspialosa volgensis), белорыбица (Stenodus leucichthys) и др.
Деление рыб на пресноводных и морских, жилых и проходных условно, ибо рыбы в биологическом отношении весьма пластичны. Есть рыбы, икра которых может одинаково успешно развиваться и в пресной и в солоноватой воде, например, ильменный пузанок (Caspialosa caspia aestuarina). Автору этой статьи приходилось в Аральском море наблюдать нерест здешней шемаи (Chalcalburnus chalcoides aralensis) и в совершенно пресной воде перед устьями Амударьи и в соленой воде посреди моря. Аральская плотва (Rutilus rutilus aralensis) и аральский сазан (Cyprinus саrріо) мечут икру по всем берегам Аральского моря (кроме, по-видимому, западного) как в совершенно пресной воде, так и в солоноватой. Равным образом и сазан, живущий перед дельтой Волги, может нереститься и в пресной воде и в солоноватой. Таким образом, пресноводные рыбы легко приспособляются к нересту в солоноватой воде. Но таких примеров, чтобы пресноводные рыбы метали икру в воде океанической солености (35‰), мы не знаем. С другой стороны, проходные рыбы без особых затруднений превращаются в жилых пресноводных. Мы уже упоминали о ладожском лососе; рыбы эта, без сомнения, произошла от проходного лосося (Salmo salar) Финского залива, но ладожский лосось никогда не уходит в море. Удавалось в прудах разводить лососей, которые размножались в пресной воде, никогда не уходя в море; между тем родители этих жилых рыб были проходными. То же известно и для нерки (Oncorhynchus nerka). Ручьевая минога, Lampetra planeri, есть пресноводное производное проходной речной миноги, L. fluviatilis, а снеток (Osmerus eperlanus morpha spirinchus) — такой же пресноводный дериват от проходной корюшки (Osmerus eperlanus). В восточной части Финского залива ряпушка (Coregonus albula) есть проходная рыба, входящая, между прочим, в Неву, тогда как во многих озерах Финляндии и северо-запада СССР это чисто пресноводная, жилая форма.
Проходные рыбы — это, очевидно, в основе обитатели пресных вод. Их пребывание в море следует рассматривать как вторичное приспособление, вызванное богатством моря кормовыми запасами. С течением времени проходные рыбы снова могут стать жилыми, как показали опыты Шо (Shaw, 1843) с кумжей (Salmo trutta) и лососем. Древнейшие известные нам рыбообразные и рыбы, оказывается, были обитателями пресных вод. Древнейшее ископаемое позвоночное, рыбообразное Astraspis (из Pteraspides), найденное в нижнесилурийских отложениях Северной Америки (Колорадо), было, по всем видимостям, обитателем пресных вод. К числу почти исключительно пресноводных относятся Acanthodii. Панцирные бесчелюстные и панцирные рыбы, находимые в изобилии в девонских отложениях типа древнего красного песчаника, в большинстве случаев принадлежат к пресноводным или солоноватоводным формам, но не к морским. Вообще, до середины девона господствовали пресноводные рыбы. Однако с середины девонского периода в морских отложениях постепенно начинает появляться все больше и больше остатков рыб, что свидетельствует в пользу образования морской фауны рыб из пресноводной. Древнейший представитель высших рыб (Actinopterygii), преобладающих в настоящее время, был найден в пресноводных отложениях, это Cheirolepis. Остатки древнейших двоякодышащих рыб (Dipnoi), в виде зубных пластинок, указываются для пресноводных отложений нижнего девона Северной Америки. Подсчеты, которые произвели Ромер и Гров (1935), показали, что в течение нижнего палеозоя число пресноводных родов в Северной Америке постепенно уменьшается, число морских — столь же постепенно растет. Это, во всяком случае, не говорит за то, чтобы палеозойские рыбы имели своей родиной море.
Одной из любопытных особенностей проходных и полупроходных рыб является наличие у них двух рас, яровой и озимой*). У лосося, или семги (Salmo salar), можно различить две формы: летнюю, или яровую, и осеннюю, или озимую. Летняя входит из моря в реки летом с хорошо развитыми половыми продуктами и мечет икру той же осенью, не подымаясь высоко вверх по реке. Напротив, осенняя (infraspecies biennis, Berg, 1932) входит в реки осенью со слабо развитыми половыми продуктами, подымается обычно высоко вверх по реке, иногда до самых верховьев, и мечет икру в следующем году осенью, проведя в реке целый год, а иногда год и несколько месяцев; все это время озимый лосось (как и вообще лосось в реке) ничем не питается.
Яровые и озимые расы выражены с большой ясностью и у осетровых. Севрюга (Acipenser stellatus), например, начинает входить из Каспийского моря в Урал в середине апреля; ход продолжается и в мае. Нерест этой севрюги — ее можно назвать яровой — происходит в мае и июне. Но с половины августа севрюга снова начинает входить в Урал и продолжает идти до половины октября; она залегает на зиму на ямы («ятови») в реке, весной подымается, идет вверх по реке и мечет икру в мае. Это, стало быть, озимая севрюга. В Дон севрюга подымается тоже дважды: весенний ход начинается вскоре по вскрытии моря и оканчивается в начале или в конце июня; разгар хода бывает в начале мая; эта севрюга — яровая — мечет икру в том же году: массовый нерест начинается во второй половине мая и продолжается до конца июня. Начало летне-осеннего хода севрюги обнаруживается слабым подъемом в июне и июле; ход увеличивается во второй половине августа и в начале сентября и продолжается в течение сентября—ноября. Эта севрюга — озимая — нерестится весной следующего года, раньше яровой, начиная с конца апреля.
*) Л. С. Берг, «Яровые и озимые расы у проходных рыб» («Известия Академии Наук», отделение мат. и ест., 1934, стр. 711-732).
Такие же две расы — яровая и озимая — имеются, помимо севрюги, и у белуги, осетра (Acipenser giildenstadti), куринского шипа (Acipenser nudiventris), у проходной стерляди из предустьевого пространства Волги.
Образованию яровых и озимых рас должно было способствовать то обстоятельство, что и у многих жилых пресноводных рыб наблюдается тенденция совершать миграции, причем у карповых, например, ясно можно видеть, что в одном и том же водоеме одни особи данного вида остаются жилыми, а другие (того же самого вида) предпринимают миграции. Даже такая, казалось бы, типично пресноводная жилая рыба, как окунь, в районе дельты Днепра весной после вскрытия реки производит передвижения из предустьевой части Днепровского лимана в Днепр и из низовых участков Днепра в более высоко расположенные участки этой реки. Осенью снова наблюдается такой же подъем окуня из лимана и дельты, причем он собирается в озерах на зимовку. Подобным образом и щука подымается из лимана в дельту Днепра два раза в год — весной и осенью.
III. Геологическое распространение рыб. Древнейшие остатки рыб известны из самых низов нижнего силура. В главконитовых песках окрестностей Петергофа встречаются мельчайшие зубы рыбы (Palaeodus, Archodus), возможно принадлежащие к Acanthodii. Из отложений нижнего силура, но более молодых, чем главконитовые пески, описаны остатки рыбообразного Astraspis из Pteraspides (Heterostraci); они встречаются в отложениях Trenton series в Колорадо. Следующие остатки рыб известны из верхнего силура (ярус лудловский и даунтонский). Они представлены обильной и разнообразной фауной рыбообразных Cephalaspides (или Osteostraci), Birkeniae (Anaspida), Pteraspides (Heterostraci), Coelolepides и настоящих рыб: Coccostei (Arthrodira; в верхнем силуре Подолии), Acanthodii. Девон — время расцвета панцирных рыбообразных: Cephalaspides, Pteraspides и таких же рыб: Pterichthyes (Antiarchi), Coccostei. За ничтожными исключениями (некоторые Coccostei) они все к концу девона вымирают. В девоне же достигают наибольшего разнообразия и Acanthodii. С верхнего девона появляются акулообразные (Cladoselache, Сlаdodus), а также химерообразные (Holocephali). Но замечательно, что тот класс рыб, который имеет в настоящее время наибольшее распространение, именно высшие рыбы (Teleostomi), обнаружен в девоне еще раньше акул и химер: из самого нижнего девона Шпицбергена и Германии известны остатки кистеперых (Crossopterygii), именно Porolepis из группы Osteolepides; как раз эта группа, куда относится, например, Eusthenopteron, из самых низов верхнего девона Канады, Шотландии и восточной Европы, наиболее близка к предкам четвероногих. Другая группа кистеперых, именно Coelacanthi, появляется в верхнем девоне. Несколько позже кистеперых, именно с середины среднего девона, становится нам известен другой подкласс высших рыб, именно Actinоpterygii, самый ранний представитель которого Cheirolepis (из Palaeoniscoidei) обнаружен в среднем древнем красном песчанике Шотландии. Ко времени же среднего (а по некоторым данным, даже нижнего) девона относится появление первых двоякодышащих (Dipnoi), именно Dipterus. Этот класс, равно как и кистеперые, достигает своего расцвета в течение девона — времени по разнообразию групп и быстроте их эволюции самого замечательного в истории рыб (и рыбообразных). Вероятно, не позже начала девона от кистеперых ответвились водные позвоночные, от которых произошли четвероногие (стегоцефалы; примитивные стегоцефалы известны из низов верхнего девона). С началом каменноугольного времени все панцирные рыбообразные и панцирные рыбы вымирают ). Большого развития достигают акулообразные, химерообразные и высшие рыбы из подкласса Actinopterygii (именно Palaeoniscoidei). Двоякодышащие и Асаnthodii начинают угасать. В конце карбона вымирают кистеперые из группы Osteolepides (Rhipidistia). В карбоне начинают появляться представители Actinopterygii, составляющие переход к высшим группам этого подкласса, именно к Holostei. Такова Теlеоpterina из верхнего карбона Северной Америки. Рыбы пермского периода, в общем, мало отличаются от карбоновых. Acanthodii в нижнепермское время вымирают. К концу перми вымирают и архаические Holocephali, чтобы в начале юры появиться в виде близких к современным формам (в общем, эволюция Holocephali пока неясна). Многочисленны примитивные Actinopterygii (т. н. Chondrostei), но в конце перми мы встречаем несомненного представителя Holostei — род Acentrophorus. В начале триаса исчезают двоякодышащие из группы Dipteri. В триасе постепенно начинают формироваться зачатки современной фауны рыб. Chondrostei распадаются на ряд отрядов. Holostei, среди них Amiiformes, приобретают значительное распространение. В верхнем триасе появляются Pholidophoriformes, близкие к Teleostei. Одновременно становятся известными и настоящие Teleostei (род Leptolepis из сельдеобразных). Весьма характерны семейства Pholidopleuridae и Saurichthyidae (последнее переходит и в лейас); оба они принадлежат к Chondrostei. Много кистеперых из семейства Coelacanthidae.
Юра. Впервые появляются осетрообразные (род Chondrosteus). С нижнего лейаса известны современные типы Holocephali (вымершие семейства Squalorajidae, Myriacanthidae); в нижней юре встречаются остатки представителей современного семейства Chimaeridae. Из Actinopterygii продолжают еще существовать Palaeoniscoidei, но сохраняются лишь единичные роды. Holostei многочисленны, равно как и Leptclepidae.
Мел. В нижнем мелу (вельд) вымирают последние Palaeoniscoidei (род Сосcolepis), и появляются представители современных семейств сельдеобразных (например, Еlоpidae, Clupeidae). В верхнем мелу Веrусіformes обычны, встречаются Perciformes и даже столь высоко организованные Teleostei, как Tetrodontiformes; много представителей современных семейств акул. Представитель кистеперых, Масrороmа, из Coelacanthi известен из верхнего мела Англии и Ливана. Многочисленны Holostei и сельдеобразные. Есть Lepidosteus. Попадаются рыбы, которых можно считать предками современных глубоководных рыб.
В третичное время фауна рыб быстро приобретает современный облик. Уже в палеоцене, имеется много современных типов, а во время нижнего эоцена (например, лютетский ярус Монте-Болька в Ломбардии) в морских отложениях можно встретить представителей почти всех современных отрядов рыб, включая и такие специализованные, как камбалы, причем, что весьма характерно, фауна носит тропический индо-тихоокеанский характер; в Монте-Болька представлено много современных родов, например, Sphyraena, Platax, Dentex, Thunnus, Lophius, Scatophagus, Diodon и мн. др. Среди вымерших родов в фауне Монте-Болька надо отметить последних представителей отряда Pycnodontiformes (из Holostei) — Pycnodus и Palaeobalistum, а также род Blochius из вымершего семейства Blochiidae (верхний мел — олигоцен), родственного современным мечам-рыбам (Xiphiidae). Известны пресноводные рыбы средней Европы из того же лютетского яруса (но из верхнего отдела его); они представлены родами Thaumaturus (из особого семейства, близкого к лососевым), Palaeoesox (из особого семейства, близкого к щукам, точнее — близкого к Umbridae), Amphiperca (близка к семейству Serranidae). Эта фауна, прекрасно описанная, представлена не теми семействами, которые сейчас обитают в Европе, но подотряды (Salmonoidei, Esocoidei, Percoidei) те же. В конце олигоцена климат Европы стал менее жарким, и в миоцене здесь появилась фауна, более близкая к современной. Отметим, что во время нижнего миоцена в Европе обитали пресноводные рыбы — Lepidosteus и Аmіа, с того времени сохранившиеся только в Северной Америке. Верхнемиоценовая морская фауна Италии, Испании, Алжира представлена современными средиземноморскими родами, но виды другие. В солоноватоводных отложениях сармата (верхний миоцен) фауна похожа на современную фауну Черного моря. Плиоценовая морская фауна рыб почти не отличается от современной. В Каспийском море тресковые (Gadus kwitkae Bog., Апшеронский п-ов) сохранялись еще в начале плиоцена (понтический ярус).
IV. Географическое распространение рыб.
А. Морские рыбы. По местообитанию морских рыб можно разделить на три категории: 1) прибрежных, 2) пелагических и 3) глубинных, или абиссальных.
1. Прибрежные рыбы населяют в главной своей массе мелководье, т. е. глубины не свыше 200 м. Они распределяются по следующим областям: а) арктическая, б) бореальная, в) тропическая, г) умеренная область южного полушария, д) антарктическая.
а) Арктическая область обнимает северо-восточную часть Баренцева моря, северного побережья Сибири и Северной Америки. Берингово море заселено бореальной фауной, и только северная часть его имеет переходную (субарктическую) фауну рыб. Арктическая область небогата рыбами; большим распространением здесь пользуются некоторые Cottidae (Myoxocephalus, Icelus, Triglops и др.), Cyclopteridae (включая и Liparidae), Agonidae, род Lycodes, затем Gymnelis viridis, из тресковых сайка (Boreogadus saida), некоторые камбалы. Юго-западная часть Баренцева моря и Белое море относятся к субарктической области (или подобласти). В субарктической области, наряду с арктическими формами, мы встречаем и бореальные, каковы: сельдь, треска, пикша, сайда (Pollachius virens), морской окунь. Большое количество арктических форм имеется и в Охотском море (здесь очень много Cottidae, Cyclopteridae, Lycodidae).
б) Бореальная, или умеренная область располагается между арктической и тропической. Она делится на две подобласти: 1) северно-атлантическую и 2) северно-тихоокеанскую; вторая, в общем, гораздо богаче. Замечательно присутствие многих амфибореальных видов, т. е. распространенных в северных частях Атлантического и Тихого океанов, но отсутствующих, полностью или частью, у берегов Сибири. Таковы, например, сельдь (Clupea harengus), треска (Gadus morhua), несколько камбал (Нірpoglossus, Limanda, Platessa и др.); все эти рыбы имеют, таким образом, прерывистое распространение. В связи с наблюдавшимся в 1919—38 гг. потеплением Арктики многие бореальные виды распространились на севере далеко за обычные пределы: до Карской губы дошли треска, лосось, сельдь (Clupea harengus pallasi; в 1936 г. была обнаружена в Обской губе); у берегов Новой Земли стала ловиться в изобилии треска, попадаются здесь теперь такие необычные для этих мест формы, как Scomberesox (1937), сайда (Роіlachius virens), пикша, макрель (все в 1936 г.). Бореальная область заключает большое количество рыб, особенно на южной окраине — в Средиземном море, на юге Японии, в Калифорнии. В этой зоне много тресковых, камбал, Labridae, сельдей, акул, скатов. В Беринговом море около 300 видов рыб, из коих свыше 90% принадлежат к тихоокеанско-бореальным, между тем как число арктических форм незначительно; здесь много Cottidae, Cyclopteridae, Blenniidae. Фауна рыб Черного моря, представленная без пресноводных рыб 135 видами, состоит из двух элементов: из черноморских автохтонов — остатков фауны верхнетретичных солоноватых озер, бывших на месте Черного и Каспийского морей, и из переселенцев из Средиземного моря. К числу первых относятся многие бычки (Gobiidae): Benthophilus, Caspiosoma, Knipowitschia, Hyrcanogobius, Benthophiloides, многие виды рода Gobius, Proterorhinus, сельди из родов Caspialosa и Clupeonella, осетровые, морской судак Lucioperca marina, Percarina. К средиземноморским переселенцам принадлежит большое количество (около ста) видов: представители родов Sprattus (кильки), Sardina (сардинки), Sardinella, Еngraulis (хамса), Anguilla (угорь), Mugil (кефаль), Serranus, Labridae, Scomber (макрель), Trigla (морской петух), Mullus (барбуля), Blennius (морские собачки), некоторые бычки и другие рыбы. Азовское море заключает обедненную фауну Черного моря. Фауна Балтийского моря тоже принадлежит к бореальной области. По мере продвижения на север и восток, в связи с все меньшей и меньшей соленостью воды этого моря, фауна его беднеет. В восточной части Финского залива, к востоку от меридиана устья Наровы, чисто морских рыб уже мало; кроме пресноводных форм, каковы щука, лещ, плотва и другие, в этой части Финского залива мы находим и проходных, каковы лосось, миноги — речная и морская, сельдь-финта, речной угорь, ряпушка, сиг, корюшка, а из морских: морскую сельдь (Clupea harengus membras) на восток до Сестрорецка, кильку (Sprattus sprattus balticus), макрель (Scomber scombrus), Myoxocephalus quadricornis и M. scorpius, речную камбалу (Pleuronectes flesus trachurus), треску (Gadus morhua callarias), морскую колюшку (Spinachia spinachia) и др. К числу реликтов ледникового периода относится в Балтийском море четырехрогий бычок Myoxocephalus quadricornis, в Арктике представленный подвидом М. quadricornis labradoricus.
Фауна Средиземного моря, прилегающих частей Атлантического океана (на север до Бискайского залива, на юг до Канарских островов), а в Тихом океане — южной части Японского моря и берегов южной Японии составляет переход к тропической области и может быть выделена в субтропическую подобласть. В южной части Японского моря (примерно к югу от линии: северная часть залива Петра Великого — пролив Цугара) встречается немало тропических форм. Так, в залив Петра Великого летом попадаются молот-рыба (Sphyrna zygaena ), двузуб (Diodon holacanthus), летучая рыба (Cypselurus agoo). Фауна рыб залива Петра Великого, состоящая примерно из 250 видов, гораздо богаче фауны Неаполитанского залива, откуда известно всего 180 видов.
в) Тропическая область может быть приблизительно отграничена годовыми изотермами температуры поверхности воды в 20°. Область эта отличается богатством фауны; особенно изобилуют рыбы коралловые рифы, дающие приют множеству ярко окрашенных видов (Chaetodontidae, Pomacentridae, Labridae, Scaridae, Acanthuridae, Tetrodontiformes и мн. др.). В пределах тропической области можно различить четыре подобласти: западно-атлантическую, восточно-атлантическую, индийско-западно-тихоокеанскую и западно-американскую; вторая и третья подобласти близки друг к другу, западно-американская же отличается почти полным отсутствием рыб, обитающих среди коралловых рифов.
Фауна рыб тихоокеанского побережья Центральной Америки показывает большое сходство с рыбами атлантического побережья этих же мест. По обе стороны Панамского перешейка имеется не менее 100 близких (викарирующих) видов. Есть много эндемичных родов, приуроченных только к атлантическим и тихоокеанским берегам Центральной Америки. Объясняется такое сходство тем, что еще в начале неогена Тихий и Атлантический океаны были соединены между собой в области Центральной Америки проливом. Но, с другой стороны, фауна рыб тропического тихоокеанского побережья Америки резко разнится от фауны островов Гавайских, Маркизовых, Паумоту. Причина заключается в том, что глубокое и лишенное островов море у западных берегов тропической Америки находится в таком состоянии уже в течение значительного, с геологической точки зрения, промежутка времени. Эта «преграда», как указывает Экман (1934, 1935), является для распространения тропической прибрежной фауны гораздо более мощным препятствием, чем какие-либо другие преграды. Сходство фауны рыб тропической части восточной Атлантики (включая и субтропическую фауну, например, Средиземного моря) с фауной индийско-западно-тихоокеанской области объясняется тем, что обе составляют наследие фауны, обитавшей некогда в океане Тетис; последний существовал еще в эоцене и частью даже в олигоцене.
г) Умеренная область южного полушария обнимает собой юг южной Африки, южный берег Австралии (на север примерно до Сиднея, 35° южной широты, и района Перта, 30° южной широты), Тасманию, Новую Зеландию, побережье Чили на юг до о-ва Чилое, побережье Аргентины (на север до 42° южной широты), о-в Тристан-да-Кунья. Фауна рыб этих мест носит, в общем, довольно разнородный характер. Но замечательно присутствие здесь многих родов, общих с бореальной областью северного полушария (явление биполярности). Таковы, например, Myxine, Cetorhinus, Scyliorhinus, Squalus, Galeus, Squatina, Raja, Clupea, Sprattus, Sardina, Sardinops, Engraulis, Conger, Zeus, Polyprion, Trigla, Sebastodes (в широком смысле слова), Careproctus, Agonidae, Merluccius, Micromesistius (из группы Gadus), Lycodes. Некоторые из этих родов могли проникнуть из северного полушария в южное по глубинам океана (например, Lophius, Careproctus, Merluccius), но большая часть распространилась в южное полушарие в ледниковое время, когда тропики испытали охлаждение (Берг, 1920). В умеренной зоне южного полушария есть, понятно, и тропические типы рыбы, но рыбы коралловых рифов здесь отсутствуют.
д) Антарктическая область обнимает берега Антарктическго материка и острова к северу от него: Южные Шетландские, Южные Оркнейские, Южные Сандвичевы, Южную Георгию, о-ва Буве, Марион, Крозе, Кергелен, Хэрд, Маккуори (к югу от Новой Зеландии). Для этой области известны 86 видов, из которых 65 приходятся на группу Notothenioidae (из Percoidei). Кроме того здесь встречаются Zoarcidae, Muraenolepidae, скаты, некоторые камбалы. Есть виды, имеющие циркумполярное распространение (например, некоторые представители семейства Nototheniidae: Pleurogramma antarcticum, некоторые виды рода Trematomus). Фауна рыб антарктической области весьма своеобразна, она состоит почти сплошь из эндемических видов и родов.
В качестве субантарктической подобласти Ригэн (1914) выделяет южную оконечность Южной Америки на север до о-ва Чилое и м. Бланко (Патагония, 47° южной широты), затем Фалькландские о-ва, крайний юг Новой Зеландии и прилегающие острова (Антиподов и др.). Подобласть эта отграничена изотермами поверхности воды примерно в 12°С на севере и 6° на юге. Для субантарктической подобласти характерно наличие ряда родов, общих с южной умеренной областью, далее присутствие семейства Galaxiidae, представителей семейства Nototheniidae и близких к ним, и ряда Zoarcidae. Имеется ряд биполярных родов: Myxine, Squalus, Raja, Clupea, Sprattus, Merluccius, Micromesistius.
2. Пелагические рыбы. Под этим именем мы понимаем здесь рыб, свойственных открытому морю и именно поверхностным слоям его до глубины примерно в 200 м. Водящиеся в толще воды на больших глубинах рыбы (т. н. батипелагические) будут рассмотрены ниже. Пелагические рыбы принадлежат к хорошим пловцам и потому имеют широкое распространение. Они свойственны преимущественно теплым морям. К пелагическим рыбам относятся многие акулы (например, такие гиганты, как Rhineodon или Cetorhinus), летучие рыбы (Exocoetus и др.), макрелеобразные (Scomber, Sarda, Thunnus и др.), солнце-рыбы (Моіа), луна-рыбы (Lampris), меч-рыбы (Xiphias). Некоторые рыбы ведут пелагический образ жизни только в личиночном состоянии, например, угри, своеобразные личинки которых известны под названием Leptocephalus.
3. Глубоководные рыбы принадлежат к двум категориям: или они живут в глубоких горизонтах водной толщи (батипелагические рыбы), или они обитают на дне океана, на больших глубинах (придонно-глубинные рыбы). Батапелагические рыбы имеют обычно космополитическое распространение, из придонно-глубинных же широко распространены лишь обитатели больших глубин. Большая часть глубоководных рыб принадлежит к батипелагическим. Область распространения, придонных глубинных животных начинается, вообще говоря, между 200 и 400 м, но настоящая глубинная фауна приурочена к глубинам свыше 1 000 м. Придонно-глубинный образ жизни ведут многие Macruridae, Zoarcidae, Cyclopteridae, некоторые Lophiiformes. Известны глубоководные семейства, которые существуют с верхнемелового времени, таковы, например, Halosauridae, Notacanthidae.
Б. Пресноводные рыбы. По распространению пресноводных (и солоноватоводных) рыб сушу можно разделить на две зоны — северную и южную. Северная, для которой характерно повсеместное распространение семейства карповых (Cyprinidae), обнимает собой Европу, Африку (без Мадагаскара), всю Азию, Малайский архипелаг к востоку до линии Уоллэса, Северную Америку на юг до границы между Мексикой и Гватемалой. Южная зона, где карповые отсутствуют, включает в себя Малайский архипелаг к востоку от линии Уоллэса, Новую Гвинею, Австралию, Новую Зеландию, тихоокеанские о-ва (Полинезию), Южную Америку (от Гватемалы к югу), Антильские о-ва, Мадагаскар.
Северная зона, состоящая из одного царства Arctogaea, разделяется на следующие области, секции и подобласти (описываются только те места, которые не рассматриваются более подробно ниже).
(I). Голарктическая (или периарктическая) область. Европа, северо-западная Африка (Марокко, Алжир, Тунис), Азия к северу от Тивериадского озера, бассейнов Евфрата и Тигра, Афганистан и Белуджистан — вне бассейна Индийского океана, далее Азия к северу от Гималаев и бассейна Амура, Северная Америка, к северу от Гватемалы. Семейство Salmonidae.
а. Циркумполярная секция. Бассейн Ледовитого моря в Европе, Азии, Америке и северная часть бассейна Тихого океана в Азии и Америке:
1. Циркумполярная подобласть.
2. Байкальская подобласть. Озеро Байкал (без его бассейна).
б. Мезевразийская секция. Европа, кроме бассейна Ледовитого моря, внутренняя Азия:
3. Средиземноморская подобласть.
4. Каспийская подобласть. Каспийское море (без впадающих в него рек).
5. Нагорно-азиатская подобласть.
в. Сонорская секция. Северная Америка кроме бассейна Ледовитого моря:
6. Миссисипская подобласть. К востоку от Скалистых гор.
7. Колорадская подобласть. К западу от Скалистых гор.
(II) . Сино-индийская область. Собственно Китай, Индокитай, Индостан, о-ва Малайского архипелага до линии Уоллэса.
8. Китайская подобласть. От южной границы Амурской переходной области (см. ниже) на юг до бассейна Сицзяна.
9. Индийская подобласть. Индокитай, Индия и острова Малайского архипелага на восток до линии Уоллэса.
(III) . Африканская область. Вся Африка, кроме северо-запада.
10. Среднеафриканская под область. Бассейны всех больших рек к югу до Замбези.
11. Восточноафриканская подобласть. От Абиссинии до устьев Замбези.
12. Южно-африканская подобласть. К югу от Замбези.
Южная зона заключает три области, распределяющиеся по двум царствам. Именно: мадагаскарская и австралийская области относятся к Notogaea, а южно-американская область — к Neogaea.
(IV) . Мадагаскарская область.
13. Мадагаскарская подобласть.
(V) . Австралийская область.
14. Австралийско-тихоокеанская подобласть.
19. Тасманийско-новозеландская подобласть.
(VI) . Южноамериканская (неотропическая) область.
16. Бразильская подобласть.
17. Патагонская подобласть.
На границе отдельных зоогеографических территорий, понятно, происходит смешение фаун, и мы наблюдаем переходные территории. Так, на границе между голарктической и сино-индийской областями располагается амурская (или манчжурская) переходная область, которая обнимает бассейн Амура и все реки, впадающие с материка в Татарский пролив и в Японское море на юг до Фузана (Корея), Сахалин, Японию от острова Хоккайдо вплоть до Киу-сиу, всю Корею, бассейны верхних течений рек Ялу и Ляо-хэ. Сюда же относятся бассейны рек Тугур и Уд, впадающих в Охотское море западнее устья Амура. Амурская переходная область разделяется на провинции:
Амурская провинция.
Приморская провинция.
Японская провинция.
Корейско-маньчжурская провинция.
Другие переходные области в Старом свете это:
Месопотамская, обнимающая бассейн Тигра и Евфрата, а также р. Куваик, на которой расположен г. Алеппо. Месопотамская область составляет переход между голарктической областью и индийской подобластью сино-индийской области. Одна месопотамская провинция.
Сирийская — переходная между голарктической и африканской областями. Она охватывает бассейн Мертвого моря и окрестности Дамаска. В бассейне Мертвого моря имеются нильские рыбы, проникшие сюда в то время, когда воды Нила доходили до Палестины. Одна сирийская провинция.
(I) . Остановимся теперь подробнее на зоогеографическом подразделении голарктической области в пределах Старого света (или на разделении Палеарктики). Здесь можно различить следующие подобласти и провинции циркумполярной (1—2) и мезевразийской (3—5) секций:
1. Циркумполярная подобласть.
Ледовитоморская провинция.
Тихоокеанская провинция.
2. Байкальская подобласть.
Байкальская провинция.
3. Средиземноморская подобласть.
Балтийская провинция.
Средиземноморская провинция.
Западно-балканская (Иллирийская) провинция.
Центрально-анатолийская провинция.
Иранская провинция.
Понто-каспийско-аральская провинция.
4. Каспийская подобласть.
Каспийская провинция.
5. Нагорно-азиатская подобласть.
Западно-монгольская провинция.
Балхашская провинция.
Таримская провинция.
Тибетская провинция.
Остановимся вкратце на характеристике вышеупомянутых зоогеографических областей, преимущественно в отношении рыбы, населяющих СССР.
Голарктической области свойственно распространение семейств Salmonidae, Thymallidae, Osmeridae, Umbridae, Dalliidae, Esocidae, Percidae, родов Petromyzon, Lampetra, Scaphirhynchus и Pseudoscaphirhynchus, Cottus, Lota, Tinea, Scardinius и мн. др.
1. Циркумполярная подобласть отличается сильным преобладанием семейства Salmonidae. Напротив, карповыми циркумполярная подобласть бедна, особенно тихоокеанская провинция. В рассматриваемой провинции есть только один эндемичный род, Dallia, имеющий притом очень ограниченное распространение — на крайнем северо-востоке Азии и в Аляске. Фауна пресноводных рыб циркумполярной подобласти остается удивительно постоянной на всем протяжении севера Европы, Азии и Америки.
Ледовитоморская провинция обнимает бассейн Ледовитого моря в Европе, Азии (кроме озера Байкала) и Америке. Представители надсемейства Salmonoidae составляют здесь около 30% всей фауны рыб. Ледовитоморскую провинцию в пределах Старого света мы делим на два округа: европейский и сибирский. Первый отличается присутствием рода Salmo, отсутствием родов Hucho, Brachymystax, Acipenser, широко распространенных в реках Сибири. Род Salmo идет на восток лишь немного далее р. Печоры (именно до р. Кары); в Печоре встречается только лосось, или семга (Salmo salar). В Сибири нет ни S. salar, ни S. trutta, ни форели (S. trutta m. fario). Представители рода Salmo появляются затем снова на Дальнем Востоке, в тихоокеанской провинции (на Камчатке и на охотском побережье). Начиная от правых притоков Енисея и далее к востоку появляется некоторая примесь американских видов; таковы: сиг-валек Соrеgonus cylindraceus, чукучан Catostomus саtostomus (Колыма), даллия Dallia pectoralis (Чукотская земля). В верхнее течение Оби проникли из нагорно-азиатской подобласти некоторые виды рода Oreoleuciscus, в бассейн верхнего Иртыша оттуда же представители рода Diptychus и подрода Diplophysa, а в бассейн Енисея — некоторые байкальские формы.
Фауна Колымы, по сравнению с ленской, обеднена: здесь нет тайменя (Hucho taimen), тугуна (Coregonus tugun), сибирской плотвы, язя, круглого карася (Carassius carassius), пескаря, щиповки, сибирского подкаменщика.
Тихоокеанская провинция обнимает бассейн р. Анадыря, Камчатку, все реки, впадающие в Тихий океан на юг почти до Амура (точнее, до реки Уд, которая относится уже к амурской провинции), затем Курильские острова и тихоокеанскую часть американской территории Аляски. Эта провинция отличается бедностью пресноводной фауны; здесь нет родов Rutilus, Perсa, Acerina, отсутствуют стерлядь (Acipenser ruthenus), сибирский осетр (А. bаегі), елец (Leuciscus leuciscus), но имеется целый ряд видов рода Oncorhynchus (кета, горбуша, чавыча, нерка, кижуч, сима), и снова появляется род Salmo (S. penshinensis, соответствующий лососю, и S. mykiss, заменяющий кумжу, S. trutta).
В Анадыре из карповых есть только гольян (Phoxinus phoxinus). Чукучан (Catostomus catostomus), свойственный Юкону, с одной стороны, и Колыме — с другой, отсутствует в Анадыре; это говорит о том, что соединение Аляски с северо-восточной Азией шло к северу от Берингова пролива. Налим Анадыря (Lota lota maculosa) принадлежит к американской форме, но рыбы эта не чуждается и солоноватой воды.
На Камчатке из жилых пресноводных рыб имеется только один хариус (Thymallus arcticus grubei n. mertensi), близкий к амурскому. Такая бедность, вообще, соответствует общему облику фауны Камчатки, не отличающейся изобилием видов.
2. Байкальская подобласть, заключающая одну байкальскую провинцию, обнимает собою только оз. Байкал, без притоков. В этом своеобразном озере имеются два эндемичных семейства рыб — Comephoridae и Cottocomephoridae. Вообще же из числа свыше трех десятков видов рыб, обитающих в Байкале, около половины оказываются эндемичными. Среди рыб Байкала нет никаких форм, которые заставляли бы выводить их из моря, — все это типичные обитатели пресных вод*).
*) Л. С. Берг, «О предполагаемых морских элементах в фауне и флоре Байкала» («Известия Академии Наук», отд. мат. и ест., 1934, стр. 303-326).
3. Средиземноморская подобласть обнимает всю Европу, кроме частей, входящих в состав циркумполярной подобласти, затем в Африке — Марокко, Алжир, Тунис, в Азии — бассейны Черного, Каспийского и Аральского морей, юг Малой Азии, Иран, кроме южной части его (бассейна Индийского океана) и Тигра. Подобласть эта отличается преобладанием сем. Cyprinidae; лососевые же отступают на задний план. Следующие роды эндемичны для этой подобласти: Caspialosa, Scardinius, Aspius, Chondrostoma, Alburnoides, Blicca, Vimba, Pelecus, Aspro, ряд родов бычков (*Gobiidae). Часть этих родов, обозначенных звездочкой, переселилась в пресную воду из солоноватых вод каспийской подобласти.
Балтийская провинция включает в себя бассейны Балтийского и Северного морей (кроме северной трети последнего), Англию, южную Шотландию, Ирландию, бассейны Ламанша и Бискайского залива, а также Альпы — независимо от бассейнов рек. Она отличается сравнительным изобилием лососевых; эндемичными являются только представители некоторых сигов и палий (Salvelinus). Если откинуть специально озерные формы сигов и палий, то фауна рыбы на всем протяжении от Рейна до Невы остается удивительно однообразной. Объяснение этого сходства заключается в том, что во время отступания великого ледника все реки, начиная от впадающих в Финский залив и кончая притоками Северного моря, были соединены между собою, образуя как бы одну общую водную систему.
Озера Ладожское и Онежское заключают несколько форм рыбы, общих с большими озерами Швеции и Северной Америки. Так, во всех упомянутых озерах живут пресноводные формы четырехрогого бычка Myoxocephalus quadricornis.
Замечательно, что в озерах Альп снова появляется циркумполярный род лососевых Salvelinus, отсутствующий к северу от Альп, но распространенный в Ирландии, Уэльсе, северной Англии, Шотландии, Скандинавии, Финляндии, на Кольском п-ове, в Карелии, в Ладожском и Онежском озерах. Кроме того в озерах Альп встречаются многочисленные сиги (Соregonus). При этом в своем распространении по Альпам ни Salvelinus, ни Coregonus совершенно не считаются с водоразделами. Такое распространение есть следствие ледникового времени, когда гидрографические связи, вследствие наличия ледникового покрова, были совсем другие, чем теперь.
Средиземноморская провинция обнимает бассейн Средиземного моря в Европе за исключением верхнего течения р. Роны (к балтийской провинции отходит и Женевское озеро), частей бассейнов рек По и Эч (Адидже), относящихся к балтийской провинции, западной части Балканского полуострова, бассейна северной части Эгейского моря и побережья Палестины. Сюда же относятся Тунис, Алжир, Марокко и реки Пиренейского п-ова, впадающие в Атлантический океан (но не в Бискайский залив; бассейн последнего относится к балтийской провинции). Провинция эта отличается бедностью своей фауны, так как больших рек здесь нет. Почти всюду распространена форель (Salmo trutta m. fario), хотя самой S. trutta в бассейне Средиземного моря в настоящее время нет.
Западно-балканская, или иллирийская провинция. К ней относятся Далмация, Албания и побережье дальше на юг вплоть до р. Ахелоя. Характерно наличие эндемичных родов Salmothymus, Pachychilon и Aulopyge. В р. Ахелое есть сом Silurus (Parasilurus) aristotelis.
Весьма замечательна фауна глубокого озера Охриды, или Охрид, относящегося к бассейну Адриатического моря. Глубина его до 286 м. Как и другие крупные западно-балканские озера, Преспа, Янина, Скутари, оно уже с середины третичного периода не стояло в связи с Эгейским морем. Что касается Адриатического моря, то в последний раз море с этой стороны доходило до оз. Охриды в среднемиоценовое время. В этом озере живет реликтовая форель Salmothymus ochridanus. Весьма богаты эндемичными формами охридские моллюски, Тrіclada, Oligochaeta; замечательная эндемичная охридская губка Ochridaspongia, обнаруживающая известное сходство с байкальскими Lubomirskiidae. Вообще, в фауне Охриды есть некоторое сходство с фауной Байкала.
Из всех провинций Европы западнобалканская наиболее своеобразна и богата эндемиками. Фауна рыбы этой провинции резко разнится от фауны рыбы восточной части Балканского полуострова*).
Центрально-анатолийская провинция обнимает замкнутые озера внутренних частей Малой Азии (Ликаония и соседние места), а также оз. Ван. Здесь много эндемичных видов. Замечательно сходство с фауной нагорно-азиатской подобласти (Берг, 1940).
Иранская провинция обнимает Иран, кроме бассейнов Каспийского моря, озера Урмии, реки Тигра и других рек, впадающих в Персидский залив и в Индийский океан; к этой же провинции относятся реки Туркмении, стекающие с Копет-дага, и реки Теджен и Мургаб, затем внутренние бассейны Ирана и Белуджистана. Здесь большим распространением пользуются роды Varicorhinus, Discognathichthys, Garra, Chalcalburnus, Cyprinion.
Понто-каспийско-аральская провинция охватывает бассейны Черного, Каспийского и Аральского морей, включая р. Чу с оз. Иссык-куль, но кроме верховьев Амударьи и Сырдарьи. К этой же провинции относятся реки Сары-су, Нура, Тургай и Иргиз. Сюда же принадлежат реки, впадающие в северную часть Эгейского моря, а также бассейн оз. Урмии. Для рассматриваемой провинции эндемичны роды: Caspiomyzon, Pseudoscaphirhynchus, Caspialosa, Clupeonella, Percarina, ряд родов из семейства Gobiidae и др. Некоторые роды понто-каспийско-аральской провинции имеют прерывистое распространение. Таковы, например, род Umbra, свойственный Днестру, Дунаю, Миссисипи, восточным штатам Северной Америки и бассейну Великих озер, Pseudoscaphirhynchus из бассейна Аральского моря, очень близкий к миссисипскому Scaphirhynchus, затем Misgurnus, обитающий в Европе и в восточной Азии, далее род белуг, Huso, свойственный восточной и южной Европе, а также бассейну Амура, наконец, миноги из подрода Eudontomyzon, распространенные в бассейне Черного моря, а затем в реках Ялу и Ляо-хэ.
*) L. S. Berg, “Übersicht der Verbreitung der Süsswasserfische Europas”, “Zoogeographica”, I, 1932.
В фауне рыб Волги и Каспийского моря есть северные элементы. К числу таковых относятся: белорыбица (Stenodus leucichthys), очень близкая к сибирской нельме; белозерская ряпушка (Coregonus sardinella vessicus), весьма близкая к беломорской; балтийская ряпушка (Coregonus albula), распространенная в некоторых озерах верховьев Волги; сибирский таймень (Hucho taimen); ручьевая минога (Lampetra planeri), пресноводное производное от речной миноги L. fluviatilis, свойственной бассейну Балтийского моря, но отсутствующей в бассейне Волги (ручьевая минога встречается в верхнем течении Волги); аналогичным образом в некоторых озерах верховьев Волги встречается снеток (Osmerus eperlanus m. spirinchus), пресноводная форма проходной корюшки (Osmerus eperlanus), обычной для рек бассейна Балтийского моря. Наконец, к той же категории северных форм надо отнести и каспийского лосося (Salmo trutta caspius), входящего в небольшом количестве в Волгу и иногда подымающегося вплоть до Камы и р. Белой (в районе г. Уфы).
Происхождение этих северных форм различное. Некоторые формы, например, ряпушка, снеток и ручьевая минога, водящиеся только в верхнем течении Волги, могли проникнуть в бассейн этой реки путем захвата Волгой верховьев рек, впадающих в Балтийское море. Другой путь проникновения северных форм шел через озерный бассейн, включавший в себя в позднеледниковое время Ладожское, Онежское озера и Белоозеро. Надо думать, что подобного рода соединения были у Волги не только с бассейном Балтийского моря, но и с бассейном Ледовитого.
4. Каспийская подобласть, заключающая одну каспийскую провинцию, обнимает лишь Каспийское море (см.) без притоков. Это гигантское соленое озеро населено настолько своеобразной фауной рыбы, моллюсков, ракообразных, червей, медуз и других беспозвоночных, что не возникает сомнения в необходимости выделения его в отдельную зоогеографическую единицу. Рыбы Каспийского моря принадлежат в значительной части к проходным, каковы осетр, севрюга, белуга, белорыбица, некоторые сельди, например, черноспинка (Caspialosa kessleri), волжская сельдь (Caspiatosa volgеnsis), но мы встречаем здесь и морские формы — атеринку (Atherina), морскую иглу (Syngnathus) и многочисленные роды Gobiidae. Все каспийские морские рыбы — и атеринка, и морская игла, и бычки — принадлежат не к строго морским формам, а к солоноватоводным.
Не исключается возможность, что солоноватоводные рыбы (особенно это справедливо для бычков, Gobiidae) остались в Каспийском море со времен верхнетретичных бассейнов, имевших солоноватую воду, подобно тому, как это происходило с многочисленными моллюсками (например, из семейства Cardiidae). К сказанному надо прибавить, что в Каспийском море распространены сельди из родов Caspialosa и Clupeonella. Род Caspialosa представлен в плиоценовых (именно в акчагыльских) отложениях Апшеронского п-ова (В. Богачев, 1933).
5. Нагорно-азиатская подобласть обнимает все внутренние, не имеющие стока к океану бассейны Центральной Азии, включая бассейн Балхаша, а также верховья рек Хильменда, Амударьи, Сырдарьи, Инда, Ганга, Брамапутры, Меконга, Янцзы-цзяна и Хуан-хэ. Фауна эта небогата, но весьма своеобразна, обладая большим количеством эндемичных родов. Она отличается наличием родов Schizothorax, Schizopygopsis, Diptychus, Aspiorhynchus, Nemachilus.
Из четырех провинций разбираемой подобласти в пределах Союза ССР имеются три: балхашская, таримская и тибетская. Для балхашской провинции весьма характерно присутствие эндемичного балхашского окуня, Perea schrenki, между тем как во всей остальной громадной области распространения рода Реrса встречается единственный вид, обыкновенный окунь, рыбы fluviatilis. Таримская провинция охватывает бассейн Тарима, а также внутренние бассейны к востоку от этой реки. В пределах СССР к таримской провинции относится часть бассейна Тарима, а также верхние течения Нарына и Карадарьи, принадлежащих к бассейну Сырдарьи.
Тибетская провинция включает в себя всю внутреннюю Азию между Кунь-лунем и Гималаями, а также верховья Хильменда, Инда, Брамапутры, Меконга, Янцзы-цзяна, Хуанхэ, Тарима, Амударьи (Памир). В пределах СССР к тибетской провинции, для которой эндемичен род Schizopygopsis, относятся воды Амударьи в пределах Памира, а также оз. Каракуль на Памире. Вне пределов СССР находится западно-монгольская провинция.
Амурская, или Маньчжурская переходная область. Амурская область включает в себя бассейн Амура, Тугура, р. Уды, а также Сахалин. В фауне рыб Амура мы находим своеобразное смешение форм ледовитоморской провинции, тихоокеанской провинции и немногочисленных эндемических видов и родов с большим числом китайских родов. Представителями ледовитоморской провинции являются роды: Lampetra, Hucho, Brachymystax, Coregonus, Thymallus, Lota. К числу северотихоокеанских, или охотских форм надо отнести род Oncorhynchus. Эндемичны роды Pseudaspius и Mesocottus. Весьма своеобразны реликтовые эндемичные виды: амурская белуга Huso dauricus и амурская щука Esox reicherti. К числу южных форм относятся змееглав (Ophiocephalus), желтощек (Elopichthys), толстолобик (Hypophthalmichthys), головешка (Perccottus) и мн. другие. Род Perccottus, свойственный Амуру и Китаю, близок к американскому Philypnus, обитателю пресных вод Мексики, Центральной Америки и Вест-Индии. Из китайских карповых можно упомянуть еще о родах Ctenopharyngodon, Mylopharyngodon, Squallobarbus, Xenocypris, нескольких китайских родах из группы пескарей, о китайских сомах из семейства Bagridae, о китайском «ерше» Siniperca.
Целый ряд видов, свойственных средиземноморской подобласти и отсутствующих в циркумполярной подобласти, снова появляется в близких или даже тождественных формах в амурской области. Таковы, например, горчак (Rhodeus sericeus), вьюн (Misgurnus), сазан (Cyprinus саrріо), белуга (Huso), сом (Silurus). Причина этого явления та, что в фауне амурской области сохранилось много реликтов — остатков фауны, обитавшей в голарктической области до наступления ледниковой эпохи и носившей более южный облик. Фауна эта в ледниковое время в Сибири или вымерла, или вынуждена была переселиться в более южные места.
К амурской провинции относятся и реки Тугур и Уда, впадающие в Охотское море западнее и севернее устья Амура. В реке Тугур встречаются Perccottus glehni и дальневосточный вьюн (Misgurnus anguillicaudatus), в реке Уде — горчак (Rhodeus sericeus) и щиповка (Cobitis taenia). На Сахалине встречаются: амурская щука (Esox reicherti), горчак, амурский язь (Leuciscus waleckii), дальневосточный вьюн, затем Mesocottus haitej, Perccottus glehni, но китайских карповых нет. По предположению Линдберга, Сахалин получил свою фауну из Амура, в ту эпоху четвертичного периода, когда Амур впадал, обогнув Сахалин, в Тихий океан где-то к северу от о-ва Хоккайдо*).
*) Г. У. Ландберг, «Фауна рыб Японского моря и история ее происхождения» («Известия Академии Наук», отд. мат. и ест., серия биол., 1937, №4, стр. 1243). – См. также А. Я. Таранец, «Пресноводные рыбы бассейна северо-западной части Японского моря» («Тр. Зоол. Инст. Акад. Наук», IV, 1936, стр. 530).
Приморская провинция обнимает реки, впадающие в Татарский пролив к югу от устья Амура и в Японское море по побережью Приморья и Кореи на юг почти до Фузана, а затем остров Иезо. Здесь сравнительно мало настоящих (не проходных) пресноводных рыб. Образование фауны приморской провинции Линдберг (I. с.) ставит в связь с существованием, в самом конце третичного периода и в начале четвертичного, обширного пресноводного озера на месте современного Японского моря; в это озеро впадали Амур, р. Суйфун вместе с речками залива Петра Великого, впадавшими в нее; р. Тумень-ула, возможно, тоже принадлежала к бассейну Суйфуна.
Переходим теперь к сонорской секции (Северная Америка). Здесь Cobitidae отсутствуют, но зато много Catostomidae; карповых (Сурrinidae) с трехрядными глоточными зубами нет, но изобилуют карповые с однорядными и двурядными зубами; есть один род, Leuciscus, — общий с Европой, а кроме того имеются роды, близкие к Rutilus, Phoxinus и Abramis. Характерны Centrarchidae, североамериканское семейство из Percoidei; к ним относятся Micropterus floridanus (обычно называемый М. salmoides, «largemouthed black bass»), M. dolomieu(«smallmouthed black bass»), солнечная рыба (Eupomotis gibbosus), ушастая солнечная рыба (Xenotis megalotis, см. ниже рис. 28), каменный окунь (Ambloplites rupestris), круглая солнечная рыба (Centrarchus macropterus). Многочисленны мелкие окуневые из подсемейства Etheostomini, эндемичного для пресных вод Северной Америки. Замечательны роды Percopsis, Columbia (см. ниже, рис. 26) и Aphredoderus из отряда Percopsiformes; Aplodinotus, пресноводный представитель морского семейства Sciaenidae; род Hyodon особого семейства сельдеобразных; род Amblyopsis — слепая пещерная рыба из семейства, принадлежащего к отряду Сурrinоdontiformes. Немало сомов из семейства Amiuridae. Совершенно особый отпечаток на фауну рыбы сонорской секции накладывает наличие здесь двух представителей Holostei, именно — Lepidosteus и Amia. Последняя (A. calva, «bowfin») широко распространена к востоку от Скалистых гор, на север до Великих озер; Lepidosteus же представлен 9 видами, из которых наиболее широко распространен L. osseus (от Великих озер до Миссисипи и Флориды); один вид встречается на Кубе, один доходит до Никарагуа. В пресных водах Северной Америки есть ряд родов, общих с Европой; таковы: Petromyzon, Lampetra, Acipenser, Salmo, Esox, Leuciscus, Lucioperca и др.; род Scaphirhynchus (Миссисипи) близок к Pseudoscaphirhynchus из бассейна Аральского моря.
(II) . СИНО-ИНДИЙСКАЯ ОБЛАСТЬ отличается отсутствием семейств, характерных для голарктической области, и присутствием значительного числа исключительно ей свойственных родов Cyprinidae, Catostomidae, Homalopteridae, Gyrinocheilidae, Cobitidae, множеством сомов, наличием Ophiocephalus, Luciocephalus, Anabantidae, Nandidae, Mastacembelidae, рода Monopterms из Symbranchidae, рода Etroplus из Cichlidae, свойственного Цейлону и южной Индии. На Суматре, Борнео и в Индокитае есть представитель семейства Osteoglossidae. На юге этой области встречаются удивительные рыбки из отряда Phallostethiformes (рис. 25; о них см. в систематической части).
(III) . АФРИКАНСКАЯ ОБЛАСТЬ отличается, помимо наличия большого количества карповых (особенно много здесь видов из группы Barbus) и сомов, присутствием многочисленных представителей семейства Cichlidae и двух семейств из подотряда Characinoidei; как известно, Cichlidae и Characinoidei свойственны также пресным водам Южной и Центральной Америки, причем надсемейство Characinoidae встречается как в Южной Америке, так и в Африке. Из эндемичных для Африки рыбы надо указать на чрезвычайно оригинальный отряд Mormyriformes, на семейство сельдеобразных Kneriidae, Pantodontidae, Phractolemidae, на Polypteridae, на род Parophiocephalus (семейство Ophiocephalidae); здесь встречаются роды Anabas, Маstacembelus, Symhranchus, Heterotis, Notopterus. Но самым удивительным членом тропической пресноводной фауны Африки является двоякодышащая рыба Protopterus. В пресноводной фауне африканской области наблюдается известное сходство с фауной Индии; обеим свойственны Anabantidae, Ophiocephalidae, Mastacembelus, Notopterus, из сомов Clarias, Heterobranchus, семейство Schilbeidae и Bagridae, из карповых Barilius и др. Это сходство объясняется тем, что еще в эоцене Индия и Африка были соединены между собой.
(IV) . МАДАГАСКАРСКАЯ ОБЛАСТЬ резко разнится отсутствием карповых. Как в Африке, здесь есть Cichlidae, но нет Characinoidei. Мадагаскар (вместе с Коморскими, Сейшельскими и Маскаренскими о-вами) беден настоящими пресноводными рыбами. Из 121 вида, найденного в пресных водах Мадагаскара и упомянутых островов, чисто пресноводных всего 34, из них Cichlidae 6 видов, сомов 3 вида, затем есть Cyprinodontidae, Serranidae, Gobiidae, Atherinidae. Несомненно, Мадагаскар заселился рыбами из Африки. Это было возможно в конце эоцена, когда в области Мозамбикского пролива произошла большая регрессия моря; тогда Мадагаскар был отделен от Африки лишь неглубокими лагунами; с другой стороны, от Мадагаскара шла цепь островов по направлению к Индии*).
*) J. Pellegrin, “La Faune ichtyologique des eaux douces de Madagascar” [Ann. Sc. Nat., zoolog. (10), XVII, 1934, p. 425-432].
(V) . АВСТРАЛИЙСКАЯ ОБЛАСТЬ. К ней относятся, кроме Австралии, Новой Гвинеи и Новой Зеландии, все острова Индо-Малайского архипелага на запад до линии, разделяющей о-ва Целебес и Борнео, а также Ломбок и Бали. Область эта отличается почти полным отсутствием настоящей пресноводной фауны (но здесь много представителей солоноватоводной фауны, переселившихся в реки). Из пресноводных рыбы надо упомянуть о двоякодышащей Neoceratodus, водящейся в Квинслэнде, и о роде Scleropages из Osteoglossidae. На юге Австралии и в Новой Зеландии встречаются представители семейства Retropinnidae и Haplochitonidae (оба из Salmonoidei) и семейства Galaxiidae, распространенного на север до Новой Каледонии. Есть также миноги.
(VI) . ЮЖНО-АМЕРИКАНСКАЯ, или НЕОТРОПИЧЕСКАЯ ОБЛАСТЬ отличается богатством своей фауны. Здесь нет карповых, но громадное количество сомовых, Characinoidei, Cichlidae. Характерны Gymnotoidei (к которым относится электрический угорь). Замечательно присутствие в Амазонке двоякодышащей Lepidosiren и костистых рыб из подотряда Osteoglossoidei (Arapaima, Osteoglossum), а также Symbranchus marmoratus. На острове Кубе мы встречаем смешанную фауну: здесь есть представители неотропических рыбы, например, Parapetenia из семейства Cichlidae, а с другой стороны — Lepidosteus, свойственный Северной и Центральной Америке. Замечательны обитающие в пещерах Кубы, в пресной воде, слепые или полуслепые рыбы Lucifuga и Stygicola (см. рис. 30), принадлежащие к морскому семейству Brotulidae (один из видов рода Brotula водится у берегов островов Вест-Индии).
V. Система рыб. Современную систему рыб надо вести от Иоганнеса Мюллера, который, считая рыб, рыбообразных и ланцетника за один класс Pisces, дал в 1844 г. такое подразделение:
Подкласс Leptocardii (отряд Amphioхіnі).
Подкласс Marsipobranchii, или Cyclostomi (отряды: миноги и миксины).
Подкласс Elasmobranchii (отряды: Plagiostomi, Holocephali).
Подкласс Ganoidei (отряды: Holostei, Chondrostei).
Подкласс Teleostei (отряды: Acanthopteri, Anacanthini, Pharyngognathi, Physostomi, Plectognathi, Lophobranchii).
Подкласс Dipnoi.
Если иметь ввиду только современных рыб, то высшие подразделения этой системы сохраняют свою силу и поныне.
Следующая система рыб, ныне живущих и ископаемых, предложена автором настоящей статьи (1940); под именем рыб в широком смысле понимаются как рыбообразные (Agnatha), так и настоящие рыбы.
I. Рыбообразные, или бесчелюстные (Agnatha), представляют особый надкласс позвоночных, резко отличный от настоящих рыб и вообще от всех остальных позвоночных. Бесчелюстные отличаются отсутствием челюстей. У всех ныне живущих, а также у многих ископаемых, полость обонятельного органа соединяется с полостью нижнего мозгового придатка (гипофиза). Жаберный скелет своеобразный, он находится кнаружи от жаберных артерий и жаберных нервов и не имеет формы отделенных друг от друга жаберных дуг, какие мы привыкли видеть у настоящих рыб. Полукружных каналов два. Хорда сохраняется в течение всей жизни. Ныне живущие рыбообразные имеют тело совершенно голое, у многих же ископаемых рыбообразных голова и передняя часть туловища были покрыты плотным костным панцирем. Рыбообразные делятся на несколько классов (знаком † обозначены ископаемые формы).
Класс I. † Cephalaspides (Osteostraci). Голова и передняя часть туловища покрыты плотным панцирем из костной ткани, обладающей костными клетками. Черепная коробка у одних состояла из хряща, облеченного перихондральным слоем кости, у других окостеневала сплошь. Одно непарное носовое отверстие наверху головы. Полость обонятельного органа сообщается с полостью гипофиза, но не сообщается с полостью глотки. Спинные корешки спинномозговых нервов не соединяются с брюшными. В голове есть электрические органы, иннервируемые ветвями n. facialis. Десять пар наружных жаберных отверстий снизу головы. Хвостовой плавник гетероцеркальный. Грудные плавники обычно есть, они покрыты чешуей и не заключают плавниковых лучей. Брюшных плавников нет. Один или два спинных плавника. Известны, начиная от верхнего силура и кончая верхним девоном.
Знаменитый шведский палеонтолог Стеншё в своем классическом труде «The Downtonian and Devonian Vertebrates of Spitzbergen» (1927) доказал, что Cephalaspides, действительно, были бесчелюстными. Стеншё обнаружил у Cephalaspides поразительное сходство с миногообразными, в частности с миногами (Petromyzones), что позволило с полной уверенностью отнести Cephalaspides к группе бесчелюстных, сопоставляя их именно с миногами, а не с миксинами.
Подкласс?† Birkeniae (Anaspida). Это — палеозойские современники Cephalaspides; они лишены головного панциря. Тело обычно покрыто рядами крупных чешуй. От 8 до 15 жаберных отверстий на боках тела. Биркении имели непарное носовое отверстие наверху головы, между, глазами, пинеальное отверстие за ним ряд наружных жаберных отверстий и гипоцеркальный хвост (т. е., ось тела продолжалась в нижнюю лопасть хвоста).
Класс II. Petromyzones (Hyperoartii; Marsipobranchii, или Cyclostomata, частью). Миноги. Тело совершенно голое. Костной ткани в скелете нет. Парных конечностей и их поясов нет. Хвост протоцеркальный. Одно непарное носовое (назогипофизное) отверстие. Назогипофизная полость не сообщается с глоткой. Жабры мешковидны, жаберные мешки у взрослых открываются в трубку, расположенную под пищеводом и спереди сообщающуюся с полостью глотки. «Язык» имеет вид своего рода поршня или бурава. Зубы роговые. Ротовой аппарат присасывательного типа. Четвертый сомит (первый заушной) снабжен нормальным миотомом. Миотомы не подразделены горизонтальной перегородкой на спинную и брюшную части. Верх мозга имеет перепончатую крышу; мозговые полости широки. Спинные корешки спинномозговых нервов не соединяются с брюшными. Есть парная «слюнная» железа. Обычно два спинных плавника (реже один). Дробление голобластическое.
Обычно миног и миксин соединяют в один класс Marsipobranchii (мешкожаберные), или Cyclostomata (круглоротые), но миноги и миксины настолько различны в своей организации, что они должны быть помещены в разные классы, как это предложено мною в 1922 г. и что доказано исследованиями Стеншё. Тогда как миноги, по своей анатомии, близки к Cephalaspides, миксины родственны Pteraspides (см. ниже).
Миноги представлены всего одним семейством Petromyzonidae (ср. миноговые), распространенным в пресных и соленых водах умеренных частей северного и южного полушарий. Миноги испытывают превращение: личинки их, известные под именем пескороек (Ammocoetes), настолько отличаются от взрослых, что раньше их описывали как особый род. У пескороек рот неприсасывательный, во рту нет роговых зубов, глаза скрыты под кожей, наружные жаберные отверстия лежат в борозде, внутренние отверстия жаберных мешков непосредственно открываются в переднюю часть кишечника. Образ жизни миног своеобразен. Размножаются они исключительно в пресной воде. Личинка живет на дне рек или озер около 4 лет, а затем превращается в молодую миножку. Ко времени нереста миноги испытывают замечательные видоизменения: зубы становятся тупыми, спинные плавники увеличиваются в высоту и сближаются до соприкосновения, у самки появляется анальный плавник (но лишенный лучей), у самца — половой сосочек.
Класс III. † Pteraspides (Heterostraci). Голова и передняя часть туловища покрыты костным панцирем, подобным тому, как у Cephalaspides, однако кость здесь лишена костных клеток. Наружного носового отверстия, по-видимому, не было, и носовая полость открывалась в ротовую полость. Электрических органов в голове нет. С каждой стороны по одному наружному жаберному отверстию. Хвостовой плавник гипоцеркальный. Других плавников нет. Эти панцирные бесчелюстные являются, насколько известно, самыми древними ископаемыми позвоночными: к этой группе относится Astraspis из нижнего силура штата Колорадо. Pteraspides известны вплоть до верхнего девона.
Подкласс? † Coelolepides (Thelodonti). Тело и голова сплошь покрыты однообразными шипиками или чешуей. Хвостовой плавник гипоцеркальный. Парных плавников нет. Спинного плавника нет. Есть анальный. Жаберные отверстия не обнаружены. Известны из верхнего силура и нижнего девона. У одних чешуя плакоидного типа, у других — скорее походит на ганоидную.
Подкласс? † Palaeospondyli. Небольшие, похожие на миног, голые рыбообразные, но с телами позвонков в форме обызвествленных (или окостеневших?) колец. Туловищные позвонки с обызвествленными верхними остистыми отростками, хвостовые с такими же верхними и нижними. Средний девон Шотландии.
Класс IV. Мухіnі (Hyperotreli; Marsipobranchii, или Cyciostomata, частью). Миксины. Как Petromyzones, но назо-гипофизная полость сообщается с полостью глотки. Ноздря на переднем конце головы. Нет никаких следов позвонков. Скелет черепа слабо развит. С каждой стороны от одного до пятнадцати наружных жаберных отверстий. Жаберных мешков 5—15 пар. Жаберная коробка зачаточна. Жаберные мешки соединяются с глоткой; есть ductus ;oesophageo-cutaneus. Приносящие сосуды на-правляются непосредственно к жаберным мешкам, а не к промежуткам между ними. Лабиринт с двумя ампулами; соответственные полукружные каналы переходят один в другой, почему обычно говорят, что у миксин только один полукружный канал. Стенки мозга толстые, мозговые полости редуцированы. Спинные корешки спинномозговых нервов соединяются с брюшными. Нет спинного плавника. Глаза скрыты под кожей и дегенерированы; глазных мышц нет. Дробление меробластическое.
В этом классе один отряд Myxiniformes с 3 семействами. Представители этого класса живут исключительно в море, присасываясь к крупным рыбам и питаясь их кровью.
Весьма замечательно наличие у Cephalaspides и Pteraspides настоящей костной ткани, как во внутреннем, так и в наружном скелете. Таким образом, кость существовала уже у силурийских бесчелюстных позвоночных, тогда как современные бесчелюстные — миноги и миксины — совершенно лишены костной ткани. Есть все основания, вместе со Стеншё, предполагать, что предки миног и миксин обладали костным скелетом, как наружным, так и внутренним, и что в процессе эволюции они утратили кость.
Надкласс Gnathostomata, челюстные. Сюда относятся настоящие рыбы и все наземные позвоночные (четырехногие, Tetrapoda). Челюстные отличаются присутствием челюстей и наличием трех полукружных каналов. Обонятельный орган не стоит в связи с гипофизом. Жаберный скелет (у взрослых или у зародышей) представлен отграниченными друг от друга жаберными дугами, которые лежат внутрь от жаберных артерий и жаберных нервов.
Нижеописываемые классы, относящиеся к настоящим рыбам (Pisies), — водные челюстные, во взрослом состоянии дышащие жабрами. Парные конечности, если они есть, не пятипалого типа. Непарные плавники поддерживаются специальным скелетом (не у всех он известен). Есть только внутреннее ухо.
Древнейшие настоящие рыбы, относящиеся к классу Acanthodii, появляются в верхнем силуре. Надкласс Pisces разделяется на следующие классы: † Pterichthyes, † Coccostei, † Acanthodii, Elasmobranchii, Holocephali, Dipnoi, Teleostomi.
Класс V. † Pterichthyes (Antiarchi). Эти девонские панцирные рыбы, со своеобразными грудными придатками, заменяющими грудные плавники, еще недавно (например, в «Палеонтологии» Вудварда, 1932) были относимы к бесчелюстным. Однако Стеншё (1931) показал, что Pterichthyes — настоящие рыбы (Pisces) с типичным для Gnathostomata челюстями. Мы выделяем эту группу в особый класс, так как трудные придатки этих рыбы не менее своеобразны, чем крылья птиц, но в отличие от передних конечностей всех остальных позвоночных имеют мышцы, покрытые наружным скелетом, — как у членистоногих, с тем, однако, основным различием, что скелет этот; костный. Кроме плотного наружного костного скелета в передних конечностях Pterichthyes имелся и внутренний скелет — хрящевой. Характеристика этого класса такова. Голова и передняя часть туловища покрыты костяным панцирем из настоящей костной ткани. Панцирь состоит из симметрично расположенных крупных пластинок. Головной щит подвижно соединяется с туловищным посредством двойного сочленения; мыщелок сочленения — на голове, а сочленовая ямка — на туловище. Глаза — на верху головы, сближены; между ними пинеальное отверстие. Одно наружное жаберное отверстие с каждой стороны. Есть жаберная крышка. Передние конечности в виде своеобразных веслообразных (по форме) придатков, покрытых таким же панцирем, как и тело; они соединяются с передней частью туловища посредством сочленения. Хвост гетероцеркальный. Один или два спинных плавника. Система каналов боковой линии в виде открытых борозд на костях наружного скелета; она в общем похожа на систему боковой линии у Elasmobranchii и Coccostei. Первичный хрящевой череп — не окостеневший и не обызвествленный; тоже и висцеральные дуги. Нет ларасфеноида. Носовые отверстия на верху головы, вплотную у переднего края глаз. Средний и верхний девон.
Класс VI. † Coccostei (Arthrodira). Эти панцирные нижнепалеозойские рыбы в известных отношениях близки к Pterichthyes (с которыми Стеншё соединяет их в группу Ріасоdermi), но лишены вышеописанных характерных грудных придатков. Стеншё обнаружил, что в нижней челюсти у Coccostei имелся мекелев хрящ, частью окостеневший. Таким образом, Coccostei, хотя и имели весьма своеобразные челюсти, принадлежали к настоящим рыбам.
Голова и передняя часть тела, как правило, покрыты костным панцирем из крупных, симметричных пластинок. Парасфеноида нет. У некоторых мекелев хрящ и palatoquadratum частью окостеневают. Верхние и нижние дуги позвонков окостеневают. Хорда сохранялась в течение всей жизни. Ребер нет. Головной панцирь, как правило, подвижно сочленен с туловищным посредством парного сочленения, причем головка сочленения — на туловищном панцире, а сочленовная ямка — на головном. Грудные плавники отсутствуют, они заменены парой неподвижных колючек. Брюшные плавники невелики. Есть спинной плавник. Есть своего рода жаберная крышка, образованная элементом нижнечелюстной дуги (postsuborbitale). Жаберное отверстие между головой и туловищным панцирем. Глаза на боках головы. Есть пинеальное отверстие. От верхнего силура до верхнего девона (или самого нижнего карбона).
Рядом с Coccostei надо поставить своеобразных девонских рыб: Stensidella, Gemuendina (которая по внешнему виду удивительно напоминает ската) и Jagorina.
Класс VII. † Acanthodii. Строение этих замечательных рыб только в самое последнее время подробно выяснено Ватсоном (Watson, 1937). Во внутреннем скелете есть настоящая кость. Есть кожные кости. Хрящевая черепная коробка или покрыта тонким слоем перихондральной кости, или окостеневает в форме нескольких перихондральных костей, разделенных хрящом. Мелкие беспорядочно разбросанные кожные кости на крыше черепа. Челюсти образованы нёбоквадратным и мекелевым хрящами; оба эти хряща окостеневают, но кожных костей на этих элементах нет. У высших форм жаберные щели покрыты кожной жаберной крышкой, поддерживаемой нижнечелюстными лучами; у низших форм такая жаберная крышка прикрывает лишь нижнюю часть жаберных щелей, вверху же жаберные щели у низших форм открываются свободно наружу, будучи каждая прикрыта своей небольшой жаберной крышкой, поддерживаемой несколькими косточками. Подъязычная дуга отделена от челюстной полной жаберной щелью, такой же величины, как и щель между подъязычной и 1-й жаберной дугой. 4 или 5 жаберных дуг. Хорда остается в течение всей жизни, но у некоторых в области позвоночника развиваются окостенения. Есть крупные отолиты. Тело покрыто чрезвычайно мелкими четырехугольными чешуями, сходными с ганоидными. Кроме хвостового плавника, все прочие впереди снабжены сильной колючкой. Хвостовой плавник гетероцеркальный. Плавники поддерживаются лучами типа ceratotrichia, т. е., как у акулообразных; плавниковые лучи слабо развиты. От верхнего силура до нижней перми.
Класс VIII. Elasmobranchil. Акулообразные. Внутренний скелет хрящевой. Кожных костей ни на черепе, ни на челюстях, ни на плечевом поясе нет. Черепная коробка никогда не окостеневает. Череп гиостилический или амфистилический. Кожа покрыта плакоидными чешуями (кожными зубами) или голая. Жаберные лепестки прикрепляются к межжаберной перегородке всей своей длиной; 5—7 жаберных дуг. Жаберных щелей 5—7 с каждой стороны, они открываются прямо наружу, жаберной крышки нет. Жаберная щель между челюстной и подъязычной дугой редуцирована. Плавательного пузыря нет. Крупных отолитов нет. Органы обоняния — в виде парных слепых мешков, каждый с одним наружным отверстием. Самцы обычно с птеригоподиями. Есть клоака. Известны от верхнего девона до настоящего времени.

Рис. 15. Акула-лисица (Alopias vulpes).
Три подкласса, из коих Cladoselachii и Xenacanthi вымершие, а третий, Selachii, заключает и ныне живущие формы. Selachii делятся на две группы: 1) акулы (см.; рис. 15), у которых, как правило, тело веретенообразное и жаберные щели на боках головы, hyomandibulare несет жаберные лучи, и 2) скаты (рис. 16), у которых тело приплюснуто и жаберные щели на нижней части головы, на hyomandibulare нет жаберных лучей. Среди акул мы различаем отряды: Heterodontiformes, Неxanchiformes, Lamniformes, Squaliformes; среди скатов — два отряда: Rajiformes и Тоrреdiniformes (ср. скаты).
Класс IX. Holocephali. Этот класс близок к Elasmobranchii, но отличается тем, что palatoquadratum, как и у Tetrapoda, слито с черепом. Одно наружное жаберное отверстие; имеется складка кожи, покрывающая 4 внутренние жаберные щели. Зубы, по крайней мере, частью, имеют вид жующих пластинок. Клоаки нет. Ребер нет. Есть pharyngohyaie. От верхнего девона до настоящего времени. Ныне живущие представлены несколькими родами (Chimaera и др.), распространенными во всех морях (ср. химеры).
Класс X. Dipnoi. Двоякодышащие (см.). Представители этого класса, известные начиная от среднего (или нижнего?) девона, в настоящее время приурочены к пресным водам Африки (Protopterus), Австралии (Ceratodus) и Южной Америки (Lepidosiren). Эти удивительные рыбы дышат и жабрами, и легкими. Palatoquadratum слито с черепом. Одно наружное жаберное отверстие, прикрытое жаберной крышкой; последняя поддерживается специальным костным скелетом. На черепе и на плечевом поясе кожные кости. Есть парасфеноид. В этом классе наблюдаются замечательные сходства в организации с четвероногими: есть легкие и соответственные видоизменения в кровеносной системе, есть намек на внутренние ноздри (хоаны), череп автостилический, парные плавники с центральной осью.
Класс XI. Teleostomi. Высшие рыбы. Palatoquadratum не слито с черепом, и челюсти, как правило, подвешены к черепу при посредстве hyomandibulare. Обе челюсти хорошо покрыты костями. На черепе кожные кости. Есть парасфеноид. Хрящевая черепная коробка более или менее окостеневает. Клоаки, как правило, нет. Одно наружное жаберное отверстие, прикрытое жаберной крышкой. Обычно есть плавательный пузырь или легкие. Есть крупные отолиты. Никогда не бывает птеригоподиев. Два подкласса: Crossopterygii и Actinopterygii.
К Crossopterygii (кистеперым) относятся ископаемые формы, известные от нижнего девона до верхнего мела, а из ныне живущих одна Latimeria, открытая в 1938 г. у берегов южной Африки (еще недавно сюда же относили ныне живущую Polypterus). Парные плавники с мясистой, покрытой чешуей осью, вдающейся в плавник и заключающей radialia. У некоторых ископаемых есть внутренние ноздри. Два спинных плавника.
К Aciinopterygii принадлежит громадное большинство ныне живущих рыб. Известны они с середины среднего девона (Cheirolepls).
Рис. 16. Скат «морской кот» (Trygon pastinaca), самка; слева — вид сверху, справа — снизу.
Парные плавники без покрытой чешуей лопасти, вдающейся в плавник; если такая лопасть имеется, она невелика и поддерживается скелетом без центральной оси. Нет внутренних ноздрей. Этот подкласс обычно делят на три группы — Chondrostei (из ныне живущих осетровые и близкие к ним формы), Ноlostei (из ныне живущих Аmіа и Lepidosteus) и Teleostei (все прочие Actinopterygii), или на две — Chondrostei и Holostei (при этом Teleostei соединяют с Holostei). Однако, если принять во внимание ископаемые формы, то группа Chondrostei нечувствительно сливается с Holostei. Условно можно принять следующие границы: 1) Chondrostei — от Роlурteriformes до Acipenseriformes, 2) Holostei — от Amiiformes до Pholidophoriformes, 3) Teleostei — Cl eiformes (семейства Lycopteridae, Leptolepidae и др.) и до конца. Мы делим Actinopterygii на следующие отряды:
Группа А. Отряд Polypteriformes (Brachiopterygii). Тело покрыто типичными ромбическими ганоидными чешуями. Хвостовой плавник симметричный. Спинной плавник состоит из многих своеобразных плавничков; каждый плавничок поддерживается одним radiale. Грудные плавники с небольшой мясистой лопастью; плавниковые лучи сидят на многочисленных radialia, которые, в свою очередь, причленяются к хрящевой пластинке и к двум удлиненным костям, сидящим на окостеневших лопатке и коракоиде. Каждый брюшной плавник, если он есть, поддерживается четырьмя radialia, сидящими на окостеневшей тазовой кости. Осевой скелет хорошо окостеневший. Череп в общем — как у Раlаеоniscoidei. Плавательный пузырь открывается в переднюю часть кишечника с брюшной стороны, ячеистый, двойной. Роды Polypterus и Calamoichthys в пресных водах Африки.
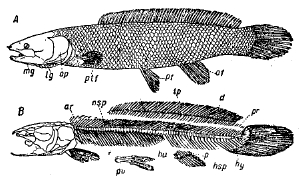
Рис. 17. Аmіа саІѵа (A) и ее скелет (В): аf - анальный плавник, аr — передние спинные radialia, d — дистальный сегмент спинного radiale, ha — гемальная дуга, hsp — гемальный отросток, hy — hypurale, lg — лучи жаберной перепонки, Ір — плавниковые лучи, mg — непарная гулярная пластинка, nsp — невральный отросток, ор — operculum, р — проксимальный сегмент radiale, рf — брюшной плавник, рr — задние спинные radialia, ptf — грудной плавник, рv — тазовая кость, r — нижнее ребро.
Группа В. Radialia грудного плавника сидят, хотя бы частью (Аmіа), непосредственно на плечевом поясе (на лопаточно-коракоидном хряще или же на лопатке и коракоиде). Сюда относится много отрядов (свыше 50), из которых мы перечислим лишь важнейшие.
Отряд † Palaeonisciformes. Маxillare тесно соединено с эктоптеригоидом и с praeoperculum. В спинном и анальном плавниках каждое radiale несет по нескольку кожных лучей. Хвостовой плавник гетероцеркальный. Чешуя типично ганоидная. От среднего девона до нижнего мела.
Триасовые отряды Redfieldiiformes (Catopteriformes), Perleidiformes и Ospiiformes составляют переход к Holostei.
Отряд † Saurichthyiformes. Замечательные рыбы, характерные для морского триаса и вымирающие в течение лейаса. Род Saurichthys (Belonorhynchus) был широко распространен: остатки его известны из Европы, Шпицбергена, Канады, Австралии. Эти крупные рыбы обнаруживают известное родство с осетровыми. Тело покрыто 4 (а не 5, как у осетровых) рядами костяных щитков или голое. МахіІІаrе, как у Palaeonisciformes. На челюстях зубы. У некоторых есть окостеневшие ребра. Хвостовой плавник симметричный.
Отряд Acipenseriformes. Тело покрыто 5 рядами жучек (Acipenseridae) или почти голое (ныне живущие Polyodontidae). Внутренняя черепная коробка остается в значительной степени хрящевой. Обе половины нёбоквадратной дуги соприкасаются по средней линии и не сочленяются с черепом. Praemaxillare слито с maxillare, а последнее тесно примыкает к palatoquadratum. Тел позвонков нет. Появляются с нижнего лейаса (Chondrosteidae). Семейство Acipenseridae (ср. ганоиды, XII, 519 сл.). Род Huso, белуги, Черное и Каспийское моря, Адриатическое, Амур; достигают веса свыше тонны. Род Acipenser, осетры; сюда относятся осетр, шип, севрюга и др.; Европа, Азия, Северная Америка. Род Scaphirhynchus, скафиринх, лопатонос; Миссисипи. Близкий род Pseudoscaphirhynchus с 3 видами в бассейне Аральского моря, семейство Polyodontidae, с очень длинным рылом. Род Polyodon; восток Северной Америки. Род Psephurus; Китай.
Отряд Amiiformes (Holostei, частью) начинает собою ряд, который раньше обозначали как Holostei. Сюда принадлежат преимущественно ископаемые рыбы, а из ныне живущих — одна североамериканская Аmіа (рис. 17). Хвостовой плавник укороченно-гетероцеркальный. Нижняя челюсть сложного строения: из dentale — spleniale, angulare, supraangulare, coronoidea (=spleniale auct.), окостенения вокруг мекелевой кости; есть gulare. Clavicula отсутствует. Maxillare свободное. Каждое radiale в спинном и анальном плавниках поддерживается одним лучом. На metapterygium сидят несколько radialia. Древнейшие представители этого отряда известны из верхней перми (Acentrophorus). Отмеченные курсивом признаки свойственны всем вышестоящим отрядам.

Рис. 18. Lepidosteus osseus.
Отряд Lepidostaiformes (Holostei , частью). Рыло сильно удлиненное, несущее на конце носовые отверстия. Praeorbitale состоит из ряда костей, несущих зубы (его обычно считают за maxillare). Помимо nasale, есть длинное praemaxillo-nasale, прободенное отверстием для обонятельного нерва. Interoperculum нет. Щеки покрыты многочисленными пластинками. Метаптеригоид сочленяется с парасфеноидом и с prooticum. Хвостовой плавник укороченно-гетероцеркальный. Позвонки вполне окостеневшие, опистоцельные. Чешуя ромбическая, ганоидная; она несет зубчики, подобные кожным зубам. Плавательный пузырь ячеистый. Один род Lepidosteus (рис. 18) в Северной и Центральной Америке и на Кубе; в Европе существовал с самого верхнего мела до нижнего миоцена.

Рис. 19. Photostomias guernei. Семейство Melanostomiatidae. Длина 60 мм. К юго-западу от Азорских островов. Грудных плавников у этого рода нет.
Отряд † Pholidophoriformes (верхний триас — верхний мел) ведет к Teleostei. И у тех, и у других нижняя челюсть простого устройства; coronoidea исчезают. Но у Pholidophoriformes хвостовой плавник укороченно-гетероцеркальный и чешуи имеют такое же строение, как у Lepidosteus.
Рис. 20. Eurypharynx bairdi. Сем. Eurypharyngidae, отряд Saccopharyngiformes. Внизу череп с громадными челюстями и подвеском, а также передние позвонки hyom – hyomandibulare, qu – quadratum, max – maxillare, mand – нижняя челюсть.
Отряд Clupeiformes. Сельдеобразные. Хвостовой плавник гомоцеркальный. Тела позвонков обычно окостеневшие (но в центре тел обычно сохраняется отверстие). Веберова аппарата нет. Есть межмышечные косточки. Верхняя челюсть обычно окаймлена как межчелюстными, так и челюстными. Мезэтмоид, обычно непарный. Как и у всех следующих отрядов, есть эндохондральное supraoccipitale, и сошник (за исключением некоторых Оsmeridae) непарный. Обычно есть мезокоракоид. Чешуя обычно циклоидная. Есть нижние и обычно верхние ребра. С верхнего триаса до настоящего времени. Как видно из диагноза, этот отряд представляет гетерогенную группу, которая со временем, без сомнения, будет расчленена на несколько самостоятельных отрядов. Приведем главнейшие подотряды.
† Leptolepldoidei. Надглазничный слизевой канал не соединяется с подглазничным. От верхнего триаса до нижнего мела.
Clupeoidel. Надглазничный слизевой канал, как и у всех следующих подотрядов и обычно отрядов, переходит на pteroticum. Есть яйцеводы. Роды Elops, Megalops, Albula, Clupea (сельди), Engraulis (анчоус или хамса) и др.
Salmonoidei. Яйцеводы недоразвиты или отсутствуют. Обычно есть жировой плавник. Роды Salmo (лосось), Oncorhynchus (кета, горбуша и др.), Stenodus (нельма, белорыбица), Coregonus (сиг), Thymallus (хариус), Osmerus (корюшка) и др.
Esocoidel. Мезэтмоид парный. Костные клетки в костях отсутствуют или имеются.
Роды Umbra, Esox (щука, E. lucius, распространена в Европе, северной Азии, Северной Америке) и др.
Stomiatoidei (рис. 19). Как Clupeoidei, но со светящимися органами. Пелагические или батипелагические рыбы.
Osteoglossoidei. Энтоптеригоид сочленяется с парасфеноидом. Носовые кости крупные, соприкасаются друг с другом. Кости крыши черепа скульптированы. Чешуя мозаична. Известны с нижнего мела. Современные живут в пресных водах Южной Америки, Австралии, Индомалайского архипелага, Таи (Сиама).
Отряд Galaxiiformes. Bulbi olfactorii прилегают к носовым капсулам. Жирового плавника нет. Яичники, как у Salmonidae. Galaxias и Neochanna: Новая Зеландия, Австралия, Новая Каледония, южная Африка, Южная Америка.
Отряд Scopeliformes. Как Clupeiformes (Stomiatoidei), но верхняя челюсть окаймлена одними praemaxiflaria. Морские рыбы, пелагические или глубоководные.
Отряд Saccopharyngiformes. Челюсти громадные, глотка растяжимая, quadratum очень велико. Костей жаберной крышки нет. Глубоководные рыбы (рис. 20).
Отряд Mormyriformes. Это один из наиболее своеобразных и резко очерченных отрядов Teleostei. Обнаруживая некоторые сходства с Clupeiformes (именно, С Notopteridae), Mormyriformes вместе с тем резко разнятся от них, равно как и от всех остальных рыб (и вообще от всех позвоночных), необыкновенно сильным развитием мозжечка. Мозжечок у них развит так, как передний мозг у человека. Глядя сверху, мы видим в мозге у Mormyrus только мозжечок, и прежние авторы рассматривали эту часть мозга у Mormyrus как передний мозг. Lobus facialis и lobus acusticus сильно развиты. Lobi olfactorii у носовых отверстий. Имеются электрические органы — производное мышечной ткани хвоста и иннервируемые посредством n. electricus, внедренного между 2-м и 3-м спинномозговыми нервами.
Bulbus arteriosus с особым отростком. Эпидермис построен своеобразно, заключая, между прочим, слои плоских горизонтальных клеток. Нёбные кости и hyomandibulare плотно прилегают к нижнему краю черепной коробки, так что намечается как бы путь к осуществлению автостилии. Есть мезокоракоид. На боках черепа большое углубление, открывающееся в полость черепа; это углубление занято круглым пузырьком, который у молодых соединяется с плавательным пузырем. Крышечные кости скрыты под кожей. Нил и пресные воды тропической Африки. Два семейства; Gymnarchidae и Mormyridae. У некоторых Mormyridae рыло сильно удлинено.
Отряд Cypriniformes. Первые четыре (а иногда пять) позвонка своеобразно видоизменены, представляя т. н. веберов аппарат, осуществляющий соединение плавательного пузыря с внутренним ухом. Как правило, открытопузырные. Обычно есть мезокоракоид. Всегда есть орбитосфеноид. Самый, обширный из всех отрядов рыбы, приуроченный в главной массе к пресным водам. Сюда относятся подотряды Characinoidei (Южная и Центральная Америка, Африка; некоторые имеют жировой плавник), Gymnotoidei (Южная и Центральная Америка; к этому подотряду принадлежит электрический «угорь», Electrophorus electricus), Cyprinoidei (карповые в широком смысле слова, множество родов в Европе, Азии, Африке и Северной Америке), Siluroidei (сомовые, см.; 28 семейств; сомы распространены повсеместно, известны с палеоцена; рис. 21).

Рис. 21. Американский сомик (Amiurus nebuloeus), длина до 45 см. Семейство Amiuridae.
Отряд Anguilliformes. Угреобразные (рис. 22). Брюшных плавников у ныне живущих нет. В плавниках нет колючек. Плавательный пузырь, если он есть, соединен с кишечником. Нет мезокоракоида. Posttemporale нет, и плечевой пояс подвешен непосредственно к позвоночнику. Praemaxillaria и мезэтмоид (а нередко и сошник) слиты в одну кость. Орбитосфеноид, если он есть, обычно парный. Позвонков много, до 260. У некоторых (например, у Cyemidae) тела позвонков в виде костяных цилиндров. Жаберные отверстия узкие. Угри, представленные более чем 20 семействами, известны с верхнего мела. Морские рыбы, некоторые глубоководные. Только один речной угорь, Anguilla anguilla, живет в реках Европы, Северной Америки, восточной Азии, но для размножения он уходит на глубины моря. У угрей из подотряда Nemichthyoidei нет окостеневшего supraoccipitale (наличие окостеневшего supraoccipitale раньше считалось характерным признаком Teleostei); у некоторых нет ни praeoperculum, ни suboperculum.

Рис. 22. Морской угорь (Conger conger).
Отряд Beloniformes. Закрытопузырные. Плавники без колючек. Брюшные плавники абдоминальные, с 6 лучами. Нижние глоточные кости вполне слиты. Верхняя челюсть окаймлена одними praemaxillaria. Преимущественно морские. Сюда относятся семейства: Belonidae (морские «щуки»; у нас имеются в Финском заливе и Черном море), Scomberesoddae (у нас в Японском и Баренцевом морях), Hemirhamphidae (некоторые, в пресной воде; у нас представители этого семейства встречаются», на Дальнем Востоке) Exocoetidae (летучие рыбы, у нас иногда на Дальнем Востоке).
Отряд Gadiformes. Трескообразные. Закрытопузырные. Плавники без колючек. Чешуя циклоидная. Брюшные плавники на горле; тазовые кости соединены с ключицей посредством связки. Хвостовой плавник симметричный. Opisthoticum очень большое, частью входит в состав стенки-лабиринта частью окаймляет отверстие для выхода n. vagus. Орбитосфеноида нет. Нет миодома и базисфеноида. Нет мезокоракоида. Обонятельные нервы не проходят через глазницу, они находятся в канале, составляющем продолжение черепной полости и расположенном над перепончатой межглазничной перегородкой. Bulbi olfactorii, как правило, у носовых капсул. Преимущественно морские рыбы, известные с палеоцена. Важнейшее в промысловом отношении семейство — тресковые (см.). Другие семейства в субтропиках, тропиках и на глубинах.

Рис. 23. Aeoliscus strigatus. Сем. Centriscidae, отр. Syngnathiformes. Индия, Япония.
Отряд Macrurif ormes. Как Gadiformes, но lobi olfactorii расположены у переднего мозга; обонятельные нервы не проходят через глазницы. Первый спинной плавник иногда с колючкой. Чешуя циклоидная или ктеноидная. Брюшные под грудными, с 5—17 лучами. Хвостовой плавник симметричный. Глубоководные рыбы.
Отряд Gasterosteiformes. Колюшкообразные. Закрытопузырные. Перед спинным плавником две или больше свободных колючек. Брюшные плавники недалеко за грудными, из одной колючки и 0—2(3) лучей. Тазовые кости не сочленяются с ключицей.
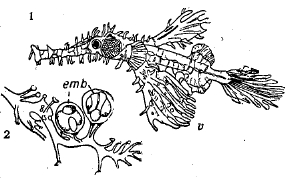
Рис. 24. Solenostomus laciniatus. Сем. Sole nostomidae, отр. Syngnathiformes; 1 — самка, v — брюшные плавники, 2 — кожный вырост выводковой камеры с зародышами (emb), расположенной у самки на брюшных плавниках.
Коракоид с эктокоракоидом. Вторая подглазничная косточка соединена с praeoperculum. Рот окаймлен одними praemaxillaria. Есть рёбра. Семейство Gasterosteidae, колюшки, в солоноватых и пресных водах северного полушария. Семейство Anlorhynchidae — северная часть Тихого океана.
Отряд Syngnathiformes. Иглообразные. Закрытопузырные. Лучи спинного (если он есть), анального и грудных плавников не ветвисты. Брюшные плавники, если есть, абдоминальные или субабдоминальные с 3—7 лучами. Тазовые кости не соединены с ключицами. Рот конечный, маленький, окаймлен или одними praemaxillaria, или же как praemaxillaria, так и maxillaria образуют край верхней челюсти. Рыло трубкообразное; сошник, мезэтмоид, quadratum и praeoperculum сильно удлинены. Ребер нет. Передние 3—6 позвонков неподвижно соединены друг с другом, кости без костных клеток. Морские рыбы, некоторые входят в пресную воду. Семейства Aulostomidae, Fistulariidae, Macrorhamphosidae, Centriscidae (рис. 23), Solenostomidae (puc. 24), Syngnathidae (морские иглы); к семейству Syngnathidae относятся и морские коньки (Hippocampus), которые встречаются и у нас в Черном море.
Рис. 25. Gulaphallus mirabilis из отряда Phallostethiformes. Самец и самка. Филиппинские острова.
Отряд Lampriformes. Закрытопузырные. Плавники без настоящих колючек. Брюшные плавники, если они есть, под грудными, с 1—17 лучами. Орбитосфеноид есть. Maxillaria выдвижные. Пелагические, частью глубинные рыбы. Сюда относятся Lampris (луна-рыба), Velifer, Trachypterus и др.
Отряд Cyprinodontiformes. Закрытопузырные. Плавники без колючек. Брюшные плавники абдоминальные, не более чем с 7 лучами. Рот окаймлен одними praemaxillaria. Нет орбитосфеноида. Нет мезокоракоида. Нет боковой линии. Один спинной плавник. Солоноватоводные и пресноводные рыбки всех тропических и умеренных частей света. Подотряд Amblyopsoidei; по большей части слепые пещерные рыбы США. Подотряд Cyprinodontoidci. Сюда относятся как яйцеродящие (Cyprinodontidae и др.), так и живородящие (Poeciliidae и др.) формы. К числу последних относится североамериканская Gambusia, пожирающая личинок комаров и в целях борьбы с малярией разведенная во многих местах (и у нас — на Кавказе и в Средней Азии).
Отряд Phallostethiformes. Удивительные рыбы пресных или солоноватых вод Индокитая, Индо-малайского архипелага и Филиппинских островов. По внешности несколько напоминают Cyprinodontiformes. Обычно два спинных плавника. Анальное отверстие под грудными плавниками или впереди их. У самцов своеобразный (единственный в своем роде среди Teleostomi) совокупительный орган, расположенный под головой; он поддерживается специальным скелетом, происходящим из первой пары ребер и из некоторых других элементов (рис. 25).
Отряд Percopsiformes. Как Реrciformes, но брюшные плавники за грудными (но не абдоминальные), с 7—8 лучами. Пресноводные рыбы Северной Америки (рис. 26). Некоторые с жировым плавником (например, Percopsis). Известны с эоцена.
Отряд Beryciformes. Как Perciformes, но есть орбитосфеноид. Брюшные плавники под грудными или субабдоминальные, с колючкой или без нее, с 3—13 мягкими лучами. Морские рыбы, известны с верхнего мела (рис. 27).
Отряд Zeiformes. Как Perciformes, но впереди анального плавника 1—4 колючки, отделенные от анального плавника промежутком. Морские рыбы. Семейство: Zeidae, Grammicolepidae, Caproidae.
Отряд Mugiliformes. Кефалеобразные. Как Perciformes, но брюшные плавники абдоминальные или субабдоминальные, и тазовые кости соединены связкой с cleithra с postcleithra. Семейство: Sphyraenidae, Mugilidae (кефали, у нас в Черном и Японском морях), Atherinidae (атеринки, у нас в Черном и Каспийском морях).
Отряд Polynemiformes. Брюшные плавники под грудными. Тазовые кости прикреплены к postcleithra. Грудные плавники состоят из двух частей, верхней и нижней; нижняя представлена несколькими нитевидными лучами. Семейство Роіуnemidae в тропических частях всех океанов.
Отряд Ophiocephaliformes. Закрытопузырные. Плавники без колючек. Нет орбитосфеноида. Рот окаймлен praemaxillaria. Брюшные плавники, если они есть, субабдоминальны; тазовые кости соединены с ключицами посредством связок. Метаптеригоид сочленяется с frontale или с sphenoticum впереди hyomandibulare. Чешуя циклоидная. Пресноводные рыбы. Два рода: Ophiocephalus в юго-восточной Азии, на севере до р. Амура, и Parophiocephalus в тропической Африке.

Рис. 26. Columbia transmontana. Семейство Реrсорsidae. Река Колумбия.
Отряд Symbranchiformes. Форма тела угреобразная. Плавательного пузыря нет. Колючек в плавниках нет. Спинной, хвостовой и анальный плавники сливаются. Брюшные плавники, если есть, на горле. Жаберные щели слиты в одну поперечную щель, расположенную на брюхе. Метаптеригоид соединяется с sphenoticum. Нет ни грудных плавников, ни лопатки, а также коракоида и грудных radialia. Жабры обычно редуцированы, дыхание происходит главным образом при посредстве кишечника. Пресные и солоноватые воды тропических частей всех материков. Род Monopterus распространен в Азии, на север до р. Ляо-хэ и острова Хондо (Япония); иногда живые экземпляры привозятся во Владивосток.
Отряд Perciformes. Окунеобразные. Закрытопузырные. В плавниках обычно есть колючие лучи. Обычно два спинных плавника. Брюшные плавники с не более чем 6 лучами, обычно под грудными (но иногда впереди грудных или немного позади их). Тазовые кости обычно прикреплены к ключицам. Глаза и череп симметричны. Нет орбитосфеноида, мезокоракоида и веберова аппарата. Мезэтмоид есть. Posttemporale обычно вильчатое. Bulbi olfactorii у переднего мозга. Хвостовой плавник с не более чем 17 главными лучами.

Maxillare обычно не окаймляет рта. Громадный отряд, населяющий преимущественно моря; известны с самого верхнего мела. Подотряды: Percoidei (рис. 28, 29), Blennioidei, Ophidioidei (рис. 30), Ammodytoidei, Callionymoidei, Siganoidei, Acanthuroidei (рис. 31), Trichiuroidei, Scombroidei, Luvaroidei, Tetragonuroidei, Stromateoidei, Anabantoidei, Luciocephaloidei, Kurtoidei (рис. 32), Ramphosoidei (ископаемый), Gobioidei (рис. 33), Cottoidei (Cathaphracti; рис. 34). У последнего 2-е infraorbitale соединено c praeoperculum.
Отряд Dactylopteriformes. Как Cottoidei, но c praeoperculum соединено 1-e infraorbitale. Nasalia слиты вместе, образуя непарную кость. Мезэтмоида нет. Posttemporale громадное. С каждой стороны пара tabularia. Первые три позвонка соединены друг с другом посредством швов. Грудные плавники громадные, разделены на две части. Семейство Dactylоpteridае с двумя родами в тропических морях. Их считают летучими рыбами, но насколько правильно, с точностью не установлено.
Отряд Thunniformes. Близки к Scombroidei, именно к семейству Cybiidae, но отличаются мощно развитой кожной сосудистой системой, связанной с сосудистым сплетением в боковых мышцах. Температура тела выше температуры воды. Известны с нижнего эоцена. Семейство Thunnidae: роды Thunnus (тунцы, у нас в Черном море и на Дальнем Востоке), Auxis и др.
Отряд Pleuronectiformes. Камбалообразные. Как Perciformes, но оба глаза на одной стороне и череп несимметричный (рис. 35). Плавники обычно без колючек. Взрослые без плавательного пузыря. Прибрежные морские рыбы, немногие входят в пресную воду. Известны с нижнего эоцена. Подотряды: Psettodoidei, в спинном и брюшных плавниках есть колючки; один род Psettodes в восточной части Атлантического океана, в Индийском и западной части Тихого. Pleuronectoidei; семейства Bothidae, Pleuronectidae, Soleidae, Cynoglossidae.
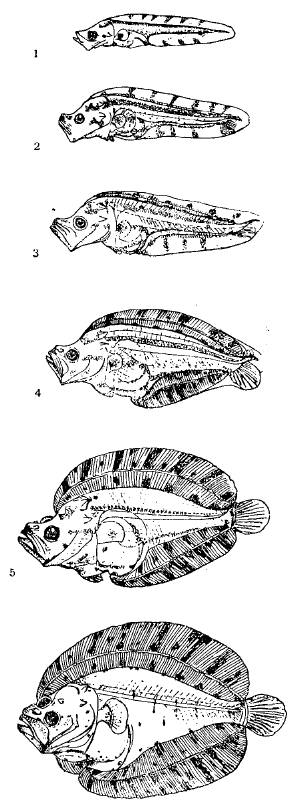
Рис. 35. Камбала Zeugopterus punctatus. Семейство Bothidae. От длины в 6 мм (№1) до длины в 12 ½ мм (№6). Сначала личинки симметричны и глаза расположены на обеих сторонах головы. Постепенно правый глаз переходит на левую сторону, и у молоди длиной 12 ½ мм оба глаза на левой стороне.
Рис. 36. Два карликовых самца Ceratias holboelli, длина 80 и 85 мм, прикрепившиеся к коже самки того же вида, длина 1 030 мм.
Отряд Mastacembeliformes. Угреобразные рыбы. Закрытопузырные. Спинной, хвостовой и анальный плавники обычно слиты. Несколько свободных колючек перед спинным плавником. В анальном плавнике три колючки. Плечевой пояс подвешен к позвоночнику. Семейство Mastacembelidae в пресных водах тропической Африки, в Евфрате, южной Азии на север до Пекина.
Отряд Echeneiformes. Прилипалообразные. Первый спинной плавник преобразован в присасывательный диск, расположенный на голове. Сем. Echeneidae, прилипалы, во всех теплых морях (и у нас в Японском).
Отряд Tetrodontiformes (Plectognathi). Как Perciformes, но posttemporaie, если оно есть, не вильчатое. Нет ни nasalia, ни infraorbitalia. Maxillaria обычно плотно соединены с praemaxillaria, иногда слиты с ними. Морские рыбы, известные с нижнего эоцена (или верхнего мела?). Подотряды Balistoidei, Ostracioidei, Tetrodontoidei, Moloidei. К последнему подотряду относится гигантская пелагическая рыба Моіа, свойственная теплым морям; летом 1936 г. добыта в заливе Петра Великого.
Отряд Gobiesociformes. Брюшные плавники видоизменены в присасывательный диск, поддерживаемый спереди ключицами, сзади — заднеключными костями. Плавательного пузыря нет. Семейство Gobiesocidae во всех океанах, у нас в Черном море.
Отряд Batrachoidiformes. Как Perciformes, но posttemporaie не вильчатое. Epioticum слито с parietale. Нет мезэтмоида. Верхняя челюсть окаймлена как посредством maxillaria, так и praemaxillaria. Брюшные плавники на горле, с 2—3 ветвистыми лучами. 4—5 грудных radialia, нижнее увеличено и на конце расширено. Семейство Batrachoididae во всех океанах.
Рис. 37. Zalises umitengu. Семейство Pegasidae. Япония, Мисаки.
Отряд Lophiiformes (Pediculati). Закрытопузырные. Первый луч первого спинного плавника, если последний имеется, расположен на голове и преобразован в своего рода «удочку». Мезэтмоид есть. Parietalia могут отсутствовать. Epiotica соприкасаются позади supraoccipitale. 2—4 radialia в грудном плавнике, нижнее увеличено и на конце расширено. Нет ни ребер, ни epipleuralia. Posttemporaie не вильчатое. Во всех морях, кроме холодных. Известны от нижнего эоцена. Подотряды Lophioidei, Antennarioidei, Ceratioidei. У некоторых семейств, принадлежащих к Ceratioidei, имеются карликовые самцы, паразитирующие на самках, — явление, единственное среди рыб (рис. 36).
Отряд Рegasiformes. Nasalia спереди слиты, образуя выдающееся, зазубренное рыло. Opisthotica, alisphenoidea, орбитосфеноид, базисфемоид, энтоптеригоид и метаптеригоид отсутствуют. Нет supracleithrum. Грудные плавники горизонтальны, с 10—18 неветвистыми лучами, при основании лучи имеют вид колючек, но дистально они делаются членистыми. Первые 6 позвонков неподвижно сочленены. Один спинной плавник. Плавательного пузыря нет, Семейство Pegasidae в тропических частях Индийского и в западной части Тихого океана (рис. 37).
Литература. Общие руководства по ихтиологии: Boulenger G. A. and Bridge Т. W., «Fishes», в «Cambrige natural history», v. VII, L., 1904; Jordan D. S., «А guide to the study of fishes», N. Y., 1905, 2 т. [главным образом систематика; сокращенное издание: «Fishes», N. Y. — L., 1925]; Goodrich E. S., «Cyclostomes and fishes», в Treatise on zoology, pt. IX, fasc. 1, L., 1909; Bronti’s, «Klassen und Ordnungen des Tierreichs», Lpz., 1901 — [изд. продолжается]; Norman J. R., «А history of fishes», L.—N. Y., 1931 [популярная]; Солдатов В. К., «Промысловая ихтиология», 2 чч., М., 1934—38; Берг Л. С., «Система рыбообразных и рыб, ныне живущих и ископаемых», Труды Зоологического института Академии наук, М., 1940, V; Суворов Е. К., «Основы общей ихтиологии», Л., 1940; Daniel J. F., «The elasmobranch fishes», 3 ed., Berkeley, California, 1934.
Общие руководства по зоологии, в которых подробно описаны рыбы: Паркер Т. Д. и Гасвелль В. А., «Руководство к зоологии», т. II, М., 1899 [пер. М. Мензбира, великолепное руководство; есть английское изд. 1940 г.]; Шимкевич В., «Курс сравнительной анатомии позвоночных животных», 3 изд., М.—Пг., 1922; Goodrich Е., «Studies on the structure and development of Vertebrates», L., 1930; «Handbuch der vergleichenden Anatomic der Wirbeltiere», hrsg. von L. Bolk, Bd. I—V, B.—Wien, 1931—39.
Основные пособия по палеонтологии рыб: Woodward A. S., «Catalogue of the fossil fishes in the British Museum», vols 1—4, L., 1889—1901; Sensiö Е. A., «Triassic fishes from Spitzbcrgen», pt. I, Vienna, 1921; pt. II, K. Svenska Vetenskaps-Akademiens handlingar, Stockholm, 1925, (3), II, № 1; его же, «The Downtonian and Devonian Vertebrates of Spitzbergen, Skrifter om Svalbard og Nordishavet, Oslo, 1927, № 12 (одно из самых замечательных произведений палеонтология, литературы); его же, «Triassic fishes from East Greenland», Meddelelser om Grönland, Köbenhavn, 1932, v. 83, № 3; его оке, «The Cephalaspids of Great Britain», L., 1933; Zitеl K. A. [and Woodward A. S.], «Text-book of palaeontology», v. II, 1932; Циттель К., «Основы палеонтологии» (русский перевод, ч. 2, печатается).
Каталоги рыбы и местные фауны. Устаревшее, но доселе незаменимое описание рыб всего света: Günthеr A., «Catalogue of the fishes in the British Museum», v. I — VIII, L., 1859—70 (переиздан без изменений в 1938 г.); Jordan D. and Evermann В., «The Fishes of North and Middle America», 4 vols, Washington, 1896—1900; Smitt F. A., «Scandinavian fishes», 2 vols, Stockholm, 1893—95; Jenkins J. T., «The fishes of the British Isles», L. — N. Y., [1925]; Moreau E., «Histoire des poissons de la France», v. I — III, P., 1881, и supplement, P., 1891; Toronese F., «Elenco dei pesci Italiani», Bolletino di pesca, 1935, IX; Buen F., de, «Catalogo de los peces ibericos», Madrid, 1935—36; Joubin L., «Faune ichtyologique de I'Atlantique Nord», Copenhague. 1929—1938; Fowler H. W., «The marine fishes of West Africa», Bulletin of the Amer. Museum of natural history, N. Y., 1936, v. LXX; Bernard K. H., «А monograph of the marine fishes of South Africa», Annals of the South African museum, Cape Town, 1925, v. XXI; Mirando Riberio A., «Fauna Brasiliensis. Peixes», Archivos do Museu Nacional de Rio de Janeiro, 1915, XVII; Pozzi A. I. у Bordale L. F., «Cuadro sistematico de los peces marinos de la Republica Argentina», Anales de la Sociedad Cientifica Argentina, Buenos Aires, 1935, t. CXX; Devinenzi G., «Peces del Uruguay», Anales del Museo de historia nacional de Montevideo, serie 2, t. I, Montevideo, 1924; Day F., «Fishes of India», pt. 1—4, L., 1875—78; Weber M. and De Beaufort L., «The fishes of the Indo-Australian Archipelago», Leidenv v. I, 1911, v. VІІ; 1936 (не окончено); Phillips W., «Bibliography of New Zealand, fishes», Wellington; 1927 (список рыб); Fowler H. W., «The fishes of Oceania»; Memoirs of the Bishop museum; Honolulu, 1928, v. X; Jordan D. S., Tanaka S. and Snyder J. O.; «A catalogue of the fishes of Japan», Journal of the College of Science University Tokyo, Tokyo, 1913, v. XXXIII, art. 1; Солдатов В. К. и Линдберг Г. У.; «Обзор рыб дальневосточник морей», Известия Тихоокеанского научного института рыбного хозяйства, т. V, Владивосток, 1930; Таранец А. Я., «Краткий определитель рыб советского Дальнего Востока и прилежащих вод», там же, 1937, т. XI; Mori T., «А revised Catalogue of the fishes of Korea», Journal Chosen Nat. Hist. Soc., № 19, 1934; Jordan D, S., Evermann В. W., Clark H. W., «Check list of the fishes and fishlike vertebrates of North and Middle America north of the northern boundary of Venezuela and Colombia», Washington, 1930; Chu Y. T., «Index piscium sinensium», Shanghai, 1931; Берг Л. C., «Рыбы пресных вод СССР и сопредельных стран», ч. 1—2, Л., 1932—33; Boulenger G. A., «Catalogue of the fresh-water fishes of Africa», v. I—IV, L., 1911—16; Eigenmann С. H., «Catalogue of the fresh-water fishes of tropical and South and Temperate America», в «Reports Of the Princeton University expeditions to Patagonia», 1896—1899, v. III, Princeton, 1905—1911; Mc Culloch A. R., «А chek-list of fishes recorded from Australia», Memoir of Australian museum, Sydney; 1929—1930, v. V.
Литература по зоогеографии морских рыб: Günther A., «An introduction to the study of fishes», Edinburgh, 1880; Ortmann A. E., «Grundzüge der marinen Tiergeographie», Jena, 1896; Jordan D. S., «А guide to the study of fishes», v. I—II, N. Y., 1905; Murray J. and Hjort J., «The depths of the ocean», L., 1912; Regan С. T., в «British Antarctic «Terra Nova» Expedition 1910», Natural; History Report, v. I—Zoology, L., 1914; Norman J. R., «The Patagonian region» (Discovery Reports, XVI, Cambridge, 1937); «The Antarctic zone», там же, v. XVIII, 1938; Hofsten N., «Die Fische des, Eisfjords; K. Svenska Vetenskaps: Akademiens Handllngar, Stockholm, 1919, v. 54, № 10; Ekman S. V., «Tiergeographie des Meeres»; Lpz., 1935.
Литература по зоогеографии пресноводных рыб: Берг Л. С., «Рыбы бассейна Амура», Записки Академии наук по физ.-мат. отд., 8 серия, т. XXIV, № 9, СПб., 1909, стр. 239—250; его же, «Рыбы пресных вод СССР и сопредельных стран», ч. 2, Л., 1933 (стр. 787— 816, карта); его же, «Разделение Палеарктики на зоогеографические области на основании распространения пресноводных рыб», Труды Первого всесоюзного географического съезда, вып. 3; Л., 1934 (Стр. 3—10, с картой); его же, «Зоогеография пресноводных рыб Передней Азии», Ученые записки Лгр. университета, № 56, 1940; Berg L. S., «Ueberslcht der Verbreitung der Süsswasser-fische Europas». Zoogeographies, Jena, 1932, Bd. I.
Прекрасный библиографический указатель: Dean В., «A bibliography of fishes», v. I—III, N. Y., 1916—23.
Л. Берг.
| Номер тома | 36 (часть 7) |
| Номер (-а) страницы | 764 |