Замораживание живых организмов
Замораживание живых организмов и оживление их после размораживания известно в науке уже давно. Так, капитан Росс (в 1821—23 гг.) во время своего путешествия по Северному Ледовитому океану подвергал гусениц бабочки Lamia действию температуры в —33°R, причем они мгновенно затвердевали и делались хрупкими, как стекло; однако, когда спустя 8 дней их переносили в теплую каюту, они вновь оживали, закукливались и давали бабочек. Подобные явления были замечены и на других животных с переменной температурой крови, как, например, у лягушек, ящериц, рыб и т. д. Так, К. Knauthe (1892) помещал лягушек в стеклянные стаканы, окруженные охлаждающей смесью снега и соли, и оставлял их на всю ночь на дворе (в апреле). Поутру их находили замерзшими и они легко разламывались; кишки оказывались промерзшими, сердце бездействовало и было окружено ледяной корой. Когда их вносили в теплую комнату, они делались около полудня мягкими, после чего начиналось слабое движение сначала левого, затем правого предсердия и, наконец, обеих сердечных камер. Первые сжатия происходили очень неправильно, а затем сердце начинало биться нормально. После этого лягушки совершенно оживали. Все эти наблюдения и опыты носили, однако, чисто случайный характер, вследствие чего не могло быть и речи о законах, которым подчинялось бы это явление. Только F. Pouchet (1866) и Н. Rödel (1886) высказали мнение, что замерзшее животное не может быть возвращено к жизни вновь, если все его соки окончательно затвердели.
С целью ближе изучить это явление, я начал в 1897 г. исследовать температуру у насекомых при помощи электрического термометра и нашел т. н. критическую точку К1, значение которой видно из следующего. Если какое-нибудь насекомое поместить в воздушную ванну, например, —20°С, то оно будет охлаждаться равномерно до некоторой точки К1, лежащей обыкновенно при —10°, но затем температура насекомого вдруг повысится до точки N (обыкновенно до —1,5°), после чего опять будет медленно падать*). Точку К1 я назвал критической, а N — началом затвердевания соков насекомого, так как соки, выжатые из насекомого, действительно начинают затвердевание при N0. Таким образом, соки насекомого от —1,5° до K1 переохлаждаются, после чего в них появляются частицы замерзшего сока, причем освобождается скрытая теплота. Следствием этого является повышение температуры до N0, которое для краткости назовем «скачком». Затем температура насекомого, по мере все большего и большего затвердевания соков, будет снова понижаться и сравняется, наконец, с температурой окружающего воздуха, т. е. будет —20°. Этот результат был проверен на нескольких ста экземпляров различных насекомых и в различных стадиях их развития.
*) Факт этот уже раньше был установлен для растений.
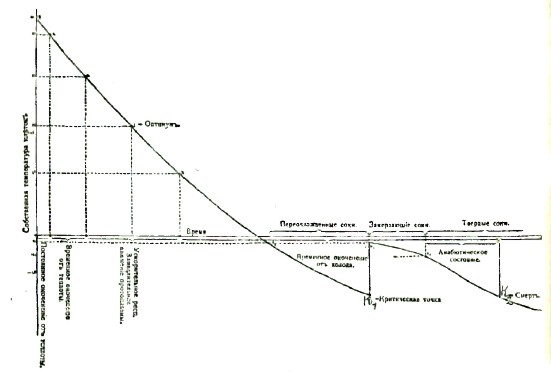
В 1898 г. я открыл мертвую точку К2, задавшись вопросом: при какой низкой температуре умирают насекомые от охлаждения? Этот вопрос можно решить, если вынимать из упомянутой холодной ванны насекомых, достигших различной температуры, и переносить их в обыкновенную температуру. Оказалось, что насекомое, вынутое из ванны перед «скачком», всегда оживало очень скоро. Насекомое, вынутое из ванны после «скачка», хотя и медленно, но все-таки оживало; но оно не могло быть более оживлено, когда его температура достигала К2 или же была ниже ее.
Таким образом, смерть насекомого не зависит от температуры, а от положения этой температуры на представленных температурных кривых, как функции времени, ибо хотя K1 равно К2, но насекомое умирает не при K1, а при К2. Поэтому-то точку К2 я и назвал мертвой точкой. Также и этот результат был проверен на нескольких ста насекомых.
В 1899 г. была изучена мною зависимость положения критической точки от скорости охлаждения, и найдено, что зависимость эта у насекомых в различных стадиях развития различна; но она подчиняется всегда одному общему правилу: при некоторой средней скорости охлаждения температура К1 достигает или максимума или минимума своей величины. Контрольные опыты с различными жидкостями, как-то: с водой, бензолом, паранитротолуолом и проч., показали то же самое. Отсюда следует, что положение точки K1 непостоянно: она может лежать и выше и ниже t° —10°, что зависит от скорости охлаждения насекомого. На положение точки K1 влияет также пол насекомого и голодовка перед опытом. Предположение, что мертвая точка К2 тоже будет изменяться параллельно с изменением точки К,, не оправдалось, так как положение точки К2, хотя при этом и изменялось, но в незначительных пределах.
В 1900—01 гг. мною была определена температура, при которой затвердевают все соки в насекомом. Калориметрические измерения показали, что соки насекомых не имеют определенной точки затвердевания, но начинают затвердевать в среднем при —1,1° (разумеется, после «скачка»), причем каждой температуре соответствует определенное количество затвердевшего сока, независимо от времени, в течение которого насекомое подвергалось действию этой температуры:
При — 1,5° их затвердевает 31%
« - 2,0 « « 73%
« - 3,0 « « 88%
« - 4 0 « « 97%
« - 4,3 « « 100%
Отсюда видно, что при —4,5° все соки в насекомом находятся в твердом виде. Таким образом, до установки анабиотического состояния у насекомых был только один шаг. В самом деле, если при —4,5° = А все соки затвердели, то циркуляция крови становится невозможной, а дыхание бесполезным, и таким образом отсутствует обмен веществ — животное более не живет, но оно и не умерло, так как не достигло мертвой точки К2. Мы должны поэтому признать, что район кривой между А и К2 представляет для насекомого анабиотическое состояние.
В 1912 г. я исследовал в этом отношении летучих мышей. В общем они дали те же результаты, как и насекомые, только критическая точка K1 была у них несколько выше, а именно, она лежала между —3° и —6°. Мертвая же точка К2 лежала около —9°. При этих исследованиях оказалось, что сперматозоиды у совокупившейся самки после размораживания и оживления животного были тоже живыми. Когда у замороженной летучей мыши, достигшей после «скачка» температуры тела —7°, была распорота грудная клетка, то ее сердце не билось и было совершенно твердым, а затем, после постепенного оттаивания, начинало биться равномерно 64 раза в минуту. В 1913 г. мною были произведены опыты над замораживанием рыб и получены те же результаты, как и с вышеупомянутыми животными.
Нужно заметить, что после размораживания животных, не достигших мертвой точки, они проявляли более или менее сильное беспокойство в течение нескольких часов и даже дней (например, карась), затем успокаивались и продолжали жить, как будто с ними ничего особенного не случилось. Продолжительность анабиотического состояния точно не была изучена; однако опыты с летучими мышами показали, что они и после 8-ми месячного пребывания в анабиозе могли быть возвращены к жизни. Обыкновенно животные возвращаются к жизни из своего анабиотического состояния после размораживания сами собой; только у некоторых экземпляров летучих мышей для этого нужно было легкое поглаживание их шерсти рукой, а у вьюна слабое раздражение переменным электрическим током.
Анабиоз животных может иметь большое значение и для практических целей. Так, обращая в анабиотическое состояние вредителей паразитов насекомых, например, наездников, живородящих мух и проч., мы имеем всегда наготове, путем их размораживания, средство — для борьбы с вредителями сельского хозяйства. Анабиоз можно было бы приложить и в пчеловодстве, чтобы пчелы не ели меда зимой; в рыбоводстве — для перевозки живой рыбы на большие расстояния, главным же образом для зернистой икры.
Литература: Zeitschr. für wiss. Zoologie, LXVI, р. 251, 1899; LXVII, р. 529, 1900; LXXI, р. 550, 1902, Biolog. Centralbl., XXI, р. 672,1901; Арх. Биол. Наук, VIIІ, р. 239, 1900. Experimentelle entomologische Studien, I (1901) и II том (1907).
П. Бахметьев.
| Номер тома | 20 |
| Номер (-а) страницы | 497 |