Животные
Животные, вместе с растениями, составляют т. н. живую, или организованную, природу, которая обыкновенно противопоставляется природе мертвой, обнимающей тела неорганические. Соответственно этому, наука, занимающаяся изучением живых существ, называется биологией в широком смысле этого слова, или наукой о жизни. Особенности живых существ выражаются в трех основных свойствах: 1) вещество, из которого состоят организмы, иначе — организованная материя, по своему химическому составу принадлежит к т. н. белковым соединениям, с примесью большого количества воды и иногда некоторого количества минеральных солей. Белковые соединения и вода, взятые вместе, являются существенной частью протоплазмы, или основного вещества всякого живого организма. 2) Живая материя постоянно изменяется, разрушаясь вследствие процессов окисления и восстановляясь вследствие поступления в нее извне нового запаса веществ, что и составляет сущность процесса обмена веществ. Конечными продуктами распадения организованного вещества являются углекислота (безазотистое соединение) и аммиак (соединение, содержащее азот). 3) Все организмы проходят последовательный цикл изменений: происходя всегда путем отделения большей или меньшей части другого организма, они приобретают со временем в большей или меньшей степени свойства последнего, размножаются в свою очередь и, наконец, умирают, т. е. перестают существовать. Различие между организмами и телами неорганическими в химическом отношении является самым важным. Хотя входящие в состав организма химические элементы все без исключения находятся также в неорганической природе, однако белковые соединения, которые построены из определенных элементов, принадлежат только организмам, являются их необходимой составной частью и в неорганических телах не встречаются совершенно. Кроме белковых соединений, в организованном веществе находятся также и другие сложные органические соединения, каковы углеводы и жиры, и более простые вещества, которые все происходят одним из двух путей — или в результате распадения белковых соединений или являясь в качестве материала для их построения.
Основные свойства животных. Будучи построены из указанных химических соединений, животные бесконечно разнообразны по химическому составу их вещества, и в этом отношении между ними нельзя найти двух тождественных. Протоплазма, образованная из этих соединений, имеет вид густой, вязкой жидкости, в которой взвешены как многочисленные твердые зернышки чрезвычайно разнообразного состава, так и крохотные капельки других жидкостей. По химической терминологии, такая смесь называется эмульсией. Пока протоплазма сохраняет хотя признак жизни, в ней происходят процессы окисления; другими словами, протоплазма находится в постоянном состоянии медленного горения, поглощая кислород извне и освобождая при своем разрушении энергию. Эта энергия и есть причина движений, в которых внешним образом выражается жизнь. С этой стороны, на живое вещество и на всех животных можно смотреть как на более или менее постоянное по форме скопление текучих частиц, притекающих, с одной стороны, и уходящих в виде продуктов окисления, с другой. Химические изменения живого вещества определяются термином метаболизм. Белковые соединения, представляющие основу протоплазмы, образованы из углерода, азота, водорода, кислорода и серы; кроме того, в теле животных постоянно встречаются фосфор, хлор, калий, натрий, магний, кальций и железо, причем некоторые из этих тел, несомненно, находятся в химическом соединении с белками. Процентный состав веществ, входящих в состав белковых соединений, изменяется в таких пределах:
Углерод от 50 до 55%
Водород от 6,5 до 7,3%
Азот от 15 до 17,6%
Кислород от 19 до 24%
Сера от 0,3 до 2,4%
Присутствие азота особенно отличает белки от углеводов и жиров, почему первые, в качестве азотистых тел, часто противополагаются двум последним группам, содержащим тела безазотистые. Нет сомнения, что величина белковых молекул очень значительна. Вследствие трудности получения белковых веществ в чистом виде, без примеси других, точное определение количества атомов, входящих в состав белковой молекулы, почти что невозможно; однако, на основании самых точных определений, можно думать, что в белковой молекуле содержится, по крайней мере, 1000 атомов. В противоположность другим телам, белковые тела не диффундируют через животные перепонки или искусственный пергамент из растворов, которые можно сделать из них в воде, и обладают способностью свертывания. Так как белковые вещества, изучаемые химией, получены из мертвых животных, они могут быть рассматриваемы в качестве продуктов распадения молекул, существовавших при жизни. Поэтому, можно думать, что белковые вещества, связанные с жизнью, существенно отличны от мертвого вещества, и, во избежание недоразумений, живые молекулы предложено называть биогенами. Процесс поглощения живой молекулой кислорода извне называется дыханием. Разрушаясь вследствие окисления, живая белковая молекула дает и такие продукты, которые уже не могут идти более на построение других живых молекул и удаляются, п. ч., накопляясь в организме, начинают вредно отражаться на его химических процессах. Процесс удаления ненужных веществ называется выделением; удаляемые продукты — экскретами, или выделениями; химические процессы, их вызывающие, — катаболизмом. Самые обыкновенные продукты разрушения, т. е. выделения, - вода, углекислота и мочевая кислота (содержит азот). Некоторые выделения, будучи вредными для организма, если остаются в протоплазме, могут косвенно приносить пользу организму, по их выведении из нее. Такие вещества называются секретами. Особые органы, выделяющие секреты, называются железами. Примером секрета можно привести желудочный сок человека и других животных. Процесс восстановления первоначальной молекулы, т. е. процесс, обратный метаболизму, носит название анаболизма или ассимиляции. Так как разрушение молекулы ведет к образованию обыкновенно более одного производного, способного к восстановлению молекулы, то можно сказать, что этот процесс, при благоприятных условиях, ведет к увеличению числа животных молекул, что внешним образом выражается в увеличении объема протоплазмы, или в росте. Восстановление живой белковой молекулы происходит на счет поступающей извне пищи, и в зависимости от того, в каком количественном отношении находятся между собой вещество, разрушающееся в организме, и вещество, поступающее в него извне, организм может увеличиваться в объеме, т. е. расти, — если количество вещества, поступающего извне, превышает количество вещества разрушающегося; не изменяться внешним образом, — если поступление вещества только уравновешивает его убыль вследствие разрушения, и постепенно разрушаться, т. е. приближаться к смерти, — если убыль вещества не покрывается его притоком извне. Так как пища, поступающая извне, должна проникать во все части протоплазмы, она должна быть растворена. Вещества, растворяющие пищу, называются ферментами и представляют собой некоторые продукты распадения живой белковой молекулы, а самый процесс перевода постороннего вещества в растворимое состояние называется пищеварением. Не переваренные или нерастворимые остатки пищи выбрасываются наружу. Движение, являющееся в результате явлений метаболизма, служит животному для разных целей, но преимущественно для добывания пищи и для того, чтобы избежать врагов. В основе способности к движению лежит способность протоплазмы изменять свой вид, быстро сокращаясь и медленно растягиваясь. При сокращении поверхность двигающейся части стремится уменьшиться, без изменения ее объема, т. е. сокращающаяся часть стремится приблизиться к сферической поверхности; напротив, при растяжении поверхность двигающейся части увеличивается. Такие части тела животного, которые в одном направлении сокращаются легче, нежели в другом, называются органами движения. Клетка, или клеточная протоплазма, обладающая специальной способностью сокращения, называется мышечной клеткой. Сокращение является в результате быстрого химического изменения или разрушения живого вещества, которое обусловливается нестойким характером живой молекулы и может быть вызвано или изменением в самом живом веществе или изменением в окружающей среде. Такое изменение действует как толчок к движению и носит название стимула; способность протоплазмы к сокращению под влиянием стимула называется раздражимостью. Стимулы различаются как внешние и внутренние, — смотря по тому, откуда исходят: из внешней среды или из самого организма. Изменение, происходящее в точке приложения стимула, часто передается далеко лежащим частям тела животных. Протоплазма, обладающая специальной способностью передачи стимулов, принадлежит нервам, образованным из отростков особых нервных клеток.
Животные и растения. Так как животные вместе с растениями образуют живую природу, которая изучается биологическими науками, необходимо, для выработки точного представления о животных выяснить различие между животными и растениями. Это различие улавливается легко только при сравнении между собой высших животных и растений. Так, животные обладают способностью произвольного движения, свободно переменяют место и принимают крайне разнообразную пищу, которая поступает через т. н. ротовое отверстие в особую пищеварительную полость. Растения, напротив, по-видимому, лишены способности произвольного движения, прикреплены неподвижно к почве и получают пищу или в жидком виде или в газах, помощью корней и листьев. Животные обладают способностью ощущений и воли, чем выражается ответ организма на внешние раздражения; растения, по-видимому, лишены этой способности, и т. д. Но, сравнивая низших животных с низшими растениями, легко видеть, что различие между ними в значительной мере сглаживается и, в конце концов, сводится к разнице в общей сумме жизненных явлений животных и растений. Между низшими животными разных групп (кораллы, гидроиды, мшанки и др.) имеются весьма многочисленные представители, которые не только лишены способности свободного передвижения и прикреплены во взрослом состоянии неподвижно на одном месте, но и по общему виду напоминают собой растения. Такие животные были известны прежде под названием зоофитов, или животно-растений. Кораллы до начала XVIII столетия считались за настоящие растения, и знаменитый Реомюр лишь с большим усилием согласился с доводами Пейсонеля, что кораллы — животные. Столь же сомнительным казалось положение в системе и других зоофитов, а между тем позднее без труда было доказано, что зоофитов, как особой группы, не существует, — что под этим именем искусственно соединены представители разных естественных групп животных, обладающие чисто внешним сходством, в результате однообразного приспособления к сходным условиям жизни. С другой стороны, среди растений находится много таких, которые обладают способностью к движению — или в смысле движения отдельных частей или в смысле даже перемещения: так называемые насекомоядные растения обладают способностью движения определенных частей в целях захватывания пищи; мимоза отвечает на механические раздражения; некоторые виды движения растений происходят под влиянием света и тепла. При этом наблюдается, что некоторые из этих движений могут быть прекращены при действии анестезирующих средств, каковы хлороформ, эфир и др. Но особенно важно в этом отношении сравнить низших животных с низшими растениями. Обширная группа амёб обнимает одноклеточные животные организмы с различной степенью дифференцировки, но характеризующиеся способностью выпускать т. н. псевдоподии (отростки, или выпячивания, тела, служащие для передвижения) и особым амёбообразным движением. Если, с другой стороны, проследить историю развития слизистых грибов (Мухоmycetes), мы найдем у них такие стадии, которые, при изучении их самих по себе, без стадий предшествующих и последующих, совершенно неотличимы от амёб — ни по строению, ни по движению. Так называемые зооспоры, или движущиеся споры, тех же грибов (стадия, за которой следует амёбообразная) и водорослей по своим движениям неотличимы от инфузорий. Сперматозоиды мхов и папоротников обладают одновременно вращательным и поступательным движениями. Наконец, на растительных, как и на животных клетках, наблюдается внутриклеточное движение протоплазмы. Следовательно, не все животные обладают движением вообще, не говоря о произвольном движении, которого лишены животные, не имеющие органов чувств и нервной системы, и не все растения неподвижны. Таким образом, движение, как критерий для различения животных от растений, отпадает. Так называемая сознательная деятельность уже потому не может служить для отличия животных от растений, что она отсутствует у множества низших животных, которые реагируют на внешние раздражения лишь постольку, поскольку чувствительна протоплазма; со стороны же чувствительности протоплазмы между животными и растениями нет разницы. Способ принятия пищи кажется хорошим критерием в указанном направлении только при поверхностном знакомстве. Среди животных существует множество паразитов, которые совершенно лишены рта и пищеварительной полости и питаются всею поверхностью своего тела, всасывая в себя жидкие питательные вещества. Таковы, например, ленточные черви. Корнеголовые (Rhizocephalidae) идут в своем приспособлении к подобному способу питания еще дальше: они забираются в тело своего животного-хозяина при помощи стебелька, который дает многочисленные корневидные разветвления, и питаются, всасывая ими жидкие питательные вещества, — совершенно так, как питаются своими корнями растения. С другой стороны, насекомоядные растения не только ловят добычу при помощи своих листьев, но и переваривают ее при помощи выделений последних. Насекомое может быть заменено кусочком мяса или круто сваренным белком. У некоторых растений на концах листьев есть особые кувшинчики, которые содержат жидкость, играющую роль пищеварительного сока, т. е. способную растворять идущие на питание твердые органические вещества. Следовательно, никак нельзя принимать в качестве общего правила, что растение питается простыми неорганическими веществами, именно углекислотой, водой и солями, а животные — только сложными органическими соединениями. Далее, мы видим, что грибы являются растениями, которые могут питаться только сложными органическими соединениями. Чужеядные растения среди высших живут также только на счет своих жертв. Наконец, есть растения, лишенные хлорофилла, тогда как есть также многие животные, несомненно, обладающие хлорофиллом. Возможно, что в основе этот хлорофилл растительного происхождения, но, так или иначе, он все-таки является принадлежащим животному организму. Процесс дыхания также общ животным и растениям. Уже давно миновало то время, когда процесс разложения углекислоты в зеленых частях растения на свете, смешивали с процессом дыхания: растение дышит, как и животное, поглощая кислород и выделяя углекислоту, но его дыхание менее энергично, нежели у животных. Наконец, процесс размножения встречается у животных и растений в одних и тех же формах — полового и бесполого размножения. Исходной точкой отправления в изучении организма животных и растений является клетка, которая в молодости одинакова у тех и других. Только с течением, времени растительная клетка, как и животная, приобретает некоторые особенности строения: у растений она более сохраняет свою индивидуальность, у животных обнаруживает большую наклонность к соединению с подобными ей для образования тканей. Однако, общая сумма жизненных явлений у животных и растений различна: в животном организме преобладают процессы окисления, распадение очень сложных органических соединений на более простые и выделение, в конечном результате процессов окисления, углекислоты и аммиака; в растении, напротив, преобладает процесс разложения углекислоты и сопровождающее его накопление углерода, т. е. преобладают процессы, обратные окислению. В результате этих различных процессов, в теле животных происходит значительная трата вещества, в разные периоды жизни лишь в той или другой степени вознаграждаемая питанием; в теле растения, напротив, наблюдается огромное накопление вещества, с избытком покрывающего убыль от процессов окисления. Так как, вообще говоря, животные неспособны сами вырабатывать сложные органические соединения, то, следовательно, для того, чтобы существовать, должны получать эти соединения извне, именно от растений, которые накопляют вещество, потребляя простые соединения и вырабатывая сложные, — это и составляет главное отличие животных от растений, устанавливая между ними вместе с тем известную зависимость.
Тот отдел биологии, который занимается изучением животных, называется зоологией. Следовательно, зоология занимается всесторонним изучением животного мира, выяснением общих законов строения, развития и жизни животных.
Изучение животных. Индивидуализированная протоплазма, или клетка, и результаты ее видоизменений могут быть изучаемы с весьма различных сторон, а именно: со стороны ее строения, места, занимаемого ею в пространстве и времени, и, наконец, со стороны деятельности тех сил, которые действуют на организм и в организме. Соответственно этому, и биология, вообще, и зоология, в частности, подразделяются на несколько отделов. Морфология изучает лишь формы живой материи, независимо от происходящих в ней процессов, и, в свою очередь, подразделяется на анатомию и гистологию, занимающиеся изучением взрослых организмов, и эмбриологию, изучающую развитие организмов. Анатомия знакомит со строением организмов, насколько оно может быть изучено без помощи микроскопа; гистология посвящена микроскопическому строению организмов, т. е. изучению клетки и клеточных комплексов. Поэтому, иначе гистология может быть определена, как учение о клетке и тканях. Иногда анатомия в обычном смысле называется макроскопической анатомией и противопоставляется гистологии, как микроскопической анатомии. Кроме того, анатомия, гистология и эмбриология могут изучать не один какой-либо объект, взятый сам по себе, а сумму объектов сравнительно, и, потому, в качестве особых предметов выделяют сравнительную анатомию, сравнительную гистологию, сравнительную эмбриологию. Тот отдел морфологии, который занимается изучением вымерших организмов, называется палеонтологией и имеет ближайшее отношение к сравнительной анатомии. Биогеография изучает современное и прошлое распространение организмов на поверхности земного шара, условия этого распространения и, потому, в свою очередь, неразрывно связана с палеонтологией. Биогеография делится на зоологическую и ботаническую географию. Наконец, физиология изучает процессы, происходящие в организме, т. е. жизненные явления организмов, которые она старается объяснить, исходя из более простых явлений неорганического мира. Так как физиология пользуется не только методом наблюдения, но и методом опытного исследования, она является наиболее точным отделом биологии и, в качестве опытной науки, может быть противопоставлена остальным, описательным, отделам биологии. Обыкновенно от физиологии отделяют биологию в узком смысле слова, которая рассматривает внешние явления жизни организма, происходящие под прямым воздействием среды и обыкновенно протекающие строго периодично. Однако, чем глубже мы будем знакомиться с собственно физиологическими процессами, тем, несомненно, теснее и теснее они будут примыкать к биологическим явлениям в узком смысле этого слова, т. е. нельзя себе представить отправления организма вне зависимости от отправления его органов. В последнее время сделаны попытки применить метод опытного исследования к морфологии, в связи с чем выделяется, в качестве особого отдела биологии, экспериментальная морфология.
Клетка. Исходной точкой отправления при изучении животного организма является клетка, под которой теперь разумеют комочек протоплазмы, содержащий внутри т. н. ядро. Протоплазма клетки образует ее тело, обладает на первый взгляд зернистым строением и, кроме ядра, позволяет различить в ней еще некоторые структуры, о которых — далее. Однако, при более тщательном изучении протоплазмы вопрос о ее строении оказывается очень сложным, и в настоящее время относительно этого существует несколько мнений. Одни считают протоплазму веществом гомогенным. Другие приписывают ей нитчатое строение, т. е. считают состоящей из нитевидных, как бы скелетных, образований и содержащегося между ними промежуточного вещества. При этом, Флеммингу эти сильно преломляющие свет ниточки, или волоконца, кажутся не соединяющимися между собой; Лейдиг и др. думают, что они образуют сеть. Видимые в протоплазме зернышки принадлежат, по этому взгляду, промежуточному веществу. Третье мнение, о так называемом пенистом строении протоплазмы, защищается Бючли (см.), который считает протоплазму состоящей из множества со всех сторон замкнутых полостей, наполненных жидким содержимым. Зернышки, или микросомы, помещаются в узловых точках, где сходятся стенки нескольких полостей. Наконец, Альтман рисует клетку состоящей из гранулей, находящихся в студневидном межгранулярном веществе. Гранулам Альтман приписывает значение элементарных организмов, называет их биобластами и приписывает им главную роль в строении плазмы, — тогда как три ранее отмеченные теории, напротив, приписывают им второстепенную роль. Протоплазма может содержать скопления жира, пигментные зернышки и пр., иногда в таком большом количестве, что тем самым совершенно скрывается ее структура. Жидкие вещества находятся в протоплазме в хорошо ограниченных полостях, т. н. вакуолях. Форма клеток чрезвычайно разнообразна: шаровидная, цилиндрическая, пирамидообразная, плоская, веретенообразная, звездчатая и т. д. Величина клеток измеряется в большинстве случаев микронами (микрон, обозначаемый μ = 0,001 mm), но колеблется в очень больших размерах. В некоторых случаях от большого количества включений и развития оболочек клетка становится непомерно большой, — например, яйцо птицы. До тех пор, пока клетка не имеет оболочки, форма ее тела может изменяться. В молодости все клетки лишены оболочки, но позднее многие из них одеваются оболочкой, которая может быть или очень тонкой или достигать значительной толщины. В последнем случае она слоиста и часто пориста. Вещество оболочки или выделяется протоплазмой клетки или представляет собой уплотненный поверхностный слой протоплазмы. Ядро при жизни клетки часто бывает невидимо, благодаря тому, что обладает одной и той же светопреломляемостью с протоплазмой. Но и в таком случае отношение ядра и протоплазмы к реактивам различно, и потому, под влиянием реактивов, ядро может быть обнаружено. По форме ядро обыкновенно шаровидно или овально; иногда — подковообразно, ветвисто и пр. Обыкновенно в каждой взрослой клетке находится только одно ядро, но в некоторых случаях число ядер бывает очень значительно. По своему составу ядро вовсе не является простым телом, т. к. в нем можно констатировать от двух до шести химически и микроскопически различных белковых веществ, из которых главные — нуклеин (хроматин), парануклеин, линин и лантанин. Из них наиболее существенны два первые. Хроматин характеризуется своей способностью краситься и по составу — присутствием фосфорной кислоты. В ядре он наблюдается в форме зернышек, нитей и сети. Парануклеин содержится в форме сильно преломляющих свет шариков, которые известны под названием ядрышек. От хроматина отличается тем, что не взбухает от прибавления воды, слабых щелочных растворов, известковой воды и пр., тогда как те же вещества заставляют хроматин взбухать и даже растворяться. Кроме того, хроматин лучше красится в кислых, парануклеин — в аммиачных растворах красок. Линин не красится обыкновенными красящими веществами и, потому, образует т. н. ахроматиновую часть ядра. Лантанин встречается иногда в виде зернышек внутри линина. Промежутки между образованиями, состоящими из нуклеина, парануклеина и линина, выполнены клеточным соком. Ядро отграничено от протоплазматического тела клетки ядерной оболочкой, которая иногда бывает ясно двуконтурна. Строение ядра весьма изменчиво, в зависимости от его состава и расположения входящих в него веществ, но особенно важно для клетки значение хроматина. Кроме ядра, в жизни клетки имеет большое значение т. н. центросома. Последняя является обыкновенно в виде одного или двух крохотных зернышек внутри протоплазмы, иногда совсем рядом с ядром и помещаясь даже в углублении на его поверхности. Вокруг центросомы в протоплазме часто бывает заметна лучистость, с лучами, сходящимися к т. н. аттракционной сфере, которая представлена светлой зоной вокруг центросомы. Клетка живет, т. е. обладает способностью к движению, раздражению, обмену веществ и размножению. Движение, по-видимому, присуще протоплазме, независимо от ядра, и представляется в разных видах. Амёбоидное движение зависит от того, что протоплазма выпускает отростки — псевдоподии, которые прикрепляются в том или другом месте и сюда подтягивают всю клетку. Псевдоподии могут быть втянуты обратно, и, благодаря их выпусканию и втягиванию, клетка может постоянно менять свои очертания. Амёбообразное движение служит как для перемещения клетки, так и для ее питания; в последнем случае псевдоподии обволакивают встречающиеся им мелкие посторонние частицы, и протоплазма клетки переваривает их, т. е. берет от них то, что может идти на ее питание. Затем, существует мерцательное движение, производимое особыми волосовидными придатками клеток — ресничками, и мышечное, присущее особенно сократимым клеткам или их видоизменениям — мышцам. Наконец, особый вид движения протоплазмы внутри клетки имеет очень важное значение для питания клетки. Раздражимостью называется способность реагировать на разные раздражения, как-то: механические, химические, термические, электрические и световые. Раздражения, смотря по их степени, могут повышать или понижать энергию жизненных явлений, и очень сильные могут привести к смерти (клетки или протоплазмы). Процесс обмена веществ, или ассимиляции, указан выше как основное свойство живого вещества. Наконец, размножение состоит в увеличении числа клеток, причем клетки происходят только путем деления других клеток. Смотря по отношению ядра к процессу деления, различают прямое, или амитотическое, деление от непрямого, или митотического. При первом способе ядро, не претерпевая существенных изменений в своем строении, перешнуровывается в определенном месте на две части, т. е. распадается на два дочерние ядра. Этот способ размножения сравнительно редок и иногда ограничивается только ядром, которое, делясь повторно, приводит к образованию многоядерных клеток. Непрямое деление — митоз, или кариокинез, — характеризуется рядом последовательных сложных изменений ядра и протоплазмы, причем, благодаря растворению оболочки ядра, между последним и протоплазмой устанавливается сложное взаимоотношение. Самым важным моментом, определяющим значение кариокинеза, является деление хроматина материнской клетки на две совершенно равные части для дочерних клеток. При этом хроматин распадается на равные отрезки — т. н. хромосомы, которые могут иметь вид петель, палочек и зерен. Число их также колеблется: их может быть 2, 4, 8,16 и больше – свыше 100. Но как вид хромосом, так и их число является характерным для клеток разных животных. Вполне равное деление хроматина на две части достигается расщеплением хромосом. Внутри протоплазмы точно также происходят очень важные явления — именно, деление центросомы, развитие вокруг нее в протоплазме лучистости, перемещение дочерних центросом к полюсам ядра и появление между ними т. н. центрального веретена. Изменения ядра и сопровождающие их явления в протоплазме происходят в известной последовательности и могут быть распределены по пяти стадиям, называемым так: 1) профаза, 2) монастер, 3) метафаза, 4) анафаза и 5) телофаза. Профаза обнимает время приготовления покоящегося ядра к делению. В ядре его более твердые составные части выступают в виде нитей, с утолщениями на их поверхности, которые потом выравниваются, а нити завиваются и образуют клубок (спирема). Сначала этот клубок довольно плотен, но потом становится более рыхлым, и в нем можно различить хромосомы в виде петель, палочек и т. д. Ядрышки (если были) всегда исчезают в это время. Центросома в начале профазы делится на две дочерние, соединенные между собой тонкими нитями, которые представляют зачаток центрального веретена: по мере того, как центросомы расходятся к полюсам ядра, находящиеся между ними сначала короткие нити становятся длиннее и выступают яснее. Затем хромосомы располагаются звездой в экваториальной плоскости (монастер), причем, если они петлевидны, то располагаются вершинами петель к центру и концами к периферии клетки, откуда, при известном положении последней, получается звездообразная фигура. На этой стадии лучистость протоплазмы образует три системы: две группы нитей идут каждая от своей центросомы к хромосомам звезды, образуя с соответствующей стороны ее род конуса; одна состоит из нитей, идущих от одной центросомы к другой; наконец, третья образована нитями, пронизывающими остальную часть клетки и расходящимися соответственно к двум центросомам, как двум центрам. Лучи последней системы, принадлежащие двум половинам клетки, пересекаются в экваториальной плоскости клетки. Метафаза начинается с расщепления хромосом, благодаря чему, вместо одной хромосомы, получаются две и, вместо простой звезды, двойная. Между дочерними петлями теперь идут нити центрального веретена, тогда как нити, ранее перекрещивавшиеся в экваториальной плоскости, не переходят более через нее. После этого начинается перешнуровывание клеточного тела в экваториальной плоскости. В стадии анафазы обе (дочерние) звезды переходят в клубкообразную структуру (диспирема), а последняя приобретает типичную структуру покоящегося ядра. В заключение ядро восстановляет свою оболочку, и в нем появляются ядрышки. Таким образом, анафаза представляет собой процесс, обратный тому, который наблюдается в профазе. В конце концов, делящаяся клетка совершенно перешнуровывается надвое, и вместо нее являются две дочерние клетки. В течение некоторого времени в них наблюдаются остатки лучистости, — на той стороне, которой одна клетка была обращена к другой, но затем все стадии центрального веретена исчезают. В течение заключительной телофазы ядро и центросома перемещаются и занимают то положение, которое присуще им в покоящейся клетке.
Клетки могут жить самостоятельно, проходя каждая свой жизненный цикл, или могут образовать группы, с определенным строением клеток и их производных, — т. н. ткани высших животных. Свободно живущие клетки или одноклеточные животные организмы весьма разнообразны со стороны своего строения и отправлений и составляют группу простейших животных (Protozoa). Все остальные животные, тело которых состоит из тканей, следовательно, животные многоклеточные, противопоставляются первым как Metazoa. Тогда как у Protozoa клетка несет все отправления, у Metazoa разные метки выполняют разные функции.
Простейшие животные (Protozoa). Из Protozoa наиболее близка к вышеописанной клетке по своей организации т. н. амёба (Amoeba); наибольшее осложнение в организации клетки представляют инфузории, но от амёбы до инфузорий можно дойти длинным рядом переходов. Амёба представляет собой протоплазматическое тело с ядром, способное выпускать псевдоподии на любом месте своей поверхности и, потому, лишенное определенной формы. Оболочки нет, но внешний слой протоплазмы, по своей однородности и относительной плотности, отличается от внутренней зернистой массы. В последней, кроме ядра, можно видеть сравнительно крупные включения разной формы, чаще всего зеленого или желтого цвета; это — непереваримые остатки пищи амёбы, т. о. микроскопически малых растений. Так как эти включения содержатся в капельках воды, эти образования называются пищевыми вакуолями. Кроме того, внутри амёбы находится вакуоля, которая, то постепенно увеличивается в объеме, то уменьшается, быстро сокращаясь. Это т. н. сократимая вакуоля, играющая роль органа выделения. Амёба питается, захватывая пищу при помощи своих псевдоподий, дышит всей своей поверхностью, выбрасывает твердые непереваримые остатки пищи на любом месте поверхности своего тела и выводит наружу ненужные вещества в растворенном виде при помощи сократимой вакуоли. Размножение происходит обыкновенно прямым делением, но иногда наблюдались амёбы с ядрами такого строения, которое обыкновенно сопровождает явления кариокинеза. Попавши в неблагоприятные условия существования, например, при высыхании окружающей ее воды, амёба одевается особой оболочкой — цистой. В таком виде амёба, подобно песчинке, может переноситься ветром с места на место. Иногда амёба, одетая цистой, распадается внутри ее на множество округлых телец, которые, выйдя из цисты после ее разрыва, становятся каждое маленькой амёбой. Такие тельца называются спорами, а процесс их образования — спорообразованием. Целый ряд близких к амёбе организмов претерпевает дальнейшее осложнение, развивая на своем теле оболочку, раковину и другие виды твердых образований из хитина, извести и кремнезема. В связи с этим, многие простейшие животные имеют огромное значение в образовании слоев земной коры (разные осадки на дне океанов, мел и пр.). Так называемые слизистые животные (Mycetozoa) замечательны тем, что у них большое число амёб, развивающихся из спор, сливается в плазмодий. Последний является в виде ярко окрашенного пятна на поверхности дубовой коры («дубильный цвет») и под микроскопом походит на огромную амёбу с ветвистыми псевдоподиями, которые иногда образуют густую сеть. Чрезвычайно многочисленные инфузории представляют целый ряд осложнений, как в своей организации, так и в своем развитии. Будучи то свободно плавающими, то сидячими, они имеют тело, одетое тонкой кутикулярной оболочкой и покрытое местами или по всей поверхности ресничками. Постоянное углубление на одном конце служит для принятия пищи и может быть названо глоткой; непереваренные остатки пищи выбрасываются, по-видимому, также через постоянное отверстие — порошицу — где-нибудь в другом месте, хотя это отверстие заметно только при проходе через него отбросов. В наружном слое протоплазмы дифференцируются сократимые (мышечные) волоконца и палочковидные образования (трехоцисты), выбрасывающиеся наружу при раздражении в виде длинных нитей, по-видимому вызывающих параличное состояние тех микроскопических организмов, с которыми соприкасаются. Вместо одного ядра, у инфузорий часто имеются два: большое (макронуклеус) и малое (микронуклеус), иногда соприкасающиеся между собой. Явления размножения достигают у инфузорий иногда большой сложности. Кроме размножения делением (поперечным и продольным), у инфузорий наблюдается еще процесс конъюгации, сопровождаемый обменом малых ядер двух конъюгирующих особей, после чего только и наступает деление. Две особи, готовые к конъюгации, прикладываются друг к другу, после чего большое ядро каждой распадается на множество кусочков, которые постепенно исчезают, вероятно идя на питание протоплазмы. Малое ядро каждой особи, делясь повторно, распадается на восемь участков, из которых семь разрушаются, а восьмой, в свою очередь, делится на два, после чего один из последних переходит через протоплазматический мостик, соединяющий две особи, в тело другой особи и здесь сливается с остающимся в ней участком принадлежащего ей малого ядра. Спустя немного, конъюгировавшие особи расходятся, и ядро каждой, образовавшееся из слития частей малых ядер двух особей, делится дважды, давая четыре ядра. После этого инфузория делится и дает две особи, каждую с двумя ядрами, из которых одно становится большим, а другое — малым. Благодаря обмену ядер между конъюгирующими особями, процесс конъюгации весьма приближается к половому размножению сложных животных. Некоторые простейшие живут паразитически, в связи с чем их организация претерпела то или другое упрощение, а размножение происходит преимущественно спорами, что обусловливает чрезвычайно быстрое увеличение числа особей. Таковы — грегарины (Sporozoa). Некоторые паразитные простейшие, как например малярийный паразит (Plasmodium malariae), проходят чрезвычайно сложный жизненный цикл, приспособившись как к дальнейшему заражению того животного, в котором они паразитируют, так и к переносу заразы с одной особи на другую. У некоторых простейших наблюдается чередование размножения двумя различными способами, что сопровождается и структурными особенностями в особях, происходящих тем или иным путем. Наилучше исследован этот процесс у Polystomella, из корненожек, и у малярийного паразита. Сущность процесса состоит в чередовании размножения простым делением и делением, сопровождаемым конъюгацией двух особей. Это явление вполне соответствует т. н. перемежающемуся размножению некоторых Metazoa.
Хотя у многих простейших не только найдена центросома, но и определено ее отношение к процессу размножения, однако в этом отношении изучение простейших отстало от изучения клетки собственно. Зато на некоторых простейших удалось доказать, что вне периода размножения центросома находится в ядре, откуда выходит при начале кариокинетических явлений.
Многие простейшие животные образуют колонии, в которых отдельные особи связаны или отростками их тела или особым выделенным ими промежуточным веществом. У одних животных особи колонии вполне одинаковы и равноценны, у других разнятся по строению и отправлениям. Такие колониальные простейшие могут быть рассматриваемы в качестве переходов от одноклеточных организмов к многоклеточным.
Яйцо, сперматозоид и оплодотворение. Все сложные животные (Metazoa) развиваются из яйца, представляющего собой клетку, которая первоначально ничем не отличается от всякой другой эмбриональной клетки и также размножается делением, давая начало новым эмбриональным клеткам. Затем такая клетка перестает делиться и быстро опережает в росте окружающие ее клетки, вырабатывая в своей протоплазме желточные крупинки. Таким образом, получается развивающееся яйцо, или яйцеклетка, становящаяся постепенно готовым яйцом, тело которого называют желтком. Прежде ядро яйца называли зародышевым пузырьком и ядрышко — зародышевым пятнышком, но теперь, после того как доказано, что яйцо представляет собой клетку, этих терминов избегают. Кроме того, в яйце найдена также центросома. Еще позднее яйцо одевается выделяемой им тонкой желточной оболочкой, а последняя, иногда значительно утолщаясь, может стать прозрачной оболочкой, которая в некоторых случаях приобретает радиальную пористость и становится радиально-пористой оболочкой. До выделения желточной оболочки яйцо усиленно питается на счет окружающих его клеток, иногда заглатывая их, подобно амёбе; после выделения оболочки питание происходит или осмотически, или, при толстой пористой оболочке, при помощи тончайших нитевидных отростков, выпускаемых телом яйца через поры оболочки. Кроме перечисленных оболочек, яйцо часто бывает одето снаружи последних оболочками, которые выделяются окружающими клетками. Таковы, например, белок и скорлупа птичьих яиц. Желток скопляется в яйце в различном количестве и распределяется различно. Тогда как протоплазма идет на построение зародыша, и потому называется образовательным желтком, желток собственно называется дейтоплазмой, или питательным желтком. Количество и способ распределения питательного желтка в теле яйца определяют способ его деления. Основная форма яйца — шаровидная. Яйцо представляет собой женский половой элемент и развивается в новый организм в огромном большинстве случаев после слития со сперматозоидом, или мужским половым элементом.
Сперматозоид, в свою очередь, представляет видоизмененную клетку; но это доказывается историей его развития, т. к. совершенно сформировавшийся сперматозоид в огромном большинстве случаев вовсе не походит на клетку. Типичная форма сперматозоида — нить, с булавовидным утолщением на одном ее конце. Утолщение, или головка, соответствует ядру клетки; нить, или хвостик, — ее видоизмененному телу. Детальное изучение сперматозоидов обнаруживает не только большое разнообразие в подробностях их строения, но также и большую сложность последнего. В промежуточной между головкой и хвостиком части находится центросома. Таким образом, как яйцо, так и сперматозоид содержат все существенные части клетки, а именно: протоплазму тела, ядро и центросому.
Процесс слития яйца с сперматозоидом называется оплодотворением яйца. Развитие яйца без оплодотворения носит название партеногенетического (девственного) развития. Оплодотворению яйца предшествует его созревание, сводящееся к редукции хроматина яйцевого ядра. Сперматозоид также содержит уменьшенное против нормы количество хроматина, но он утрачивает часть его при своем развитии. Редукция хроматина яйцевого ядра происходит в большинстве случаев до соединения яйца с сперматозоидом, но иногда наступает и позднее. У Physa fontinalis, из моллюсков, процесс созревания и оплодотворения яйца с необычайной ясностью протекает следующим образом. Сперматозоид проникает в яйцо до его созревания, но пока последнее не наступит, остается в теле яйца видимым со всеми своими частями, т. е. головкой, центросомой и хвостиком. Созревание яйца сводится к его повторному кариокинетическому делению на две неравные части. Как и при всяком делении клетки, этот процесс начинается с распадения ядра на хромосомы, при одновременном появлении центросом и лучистости вокруг них. Вскоре вся эта кариокинетическая фигура передвигается к поверхности клетки, вместе с чем простая звезда распадается на две дочерних. На поверхности яйца образуется бугорок, где помещается одна центросома, одна дочерняя звезда и половина веретена, после чего бугорок отделяется от яйца, образуя первый направляющий пузырек. Затем образуется второй направляющий пузырек, но тогда как при образовании первого дочерние звезды получаются путем расщепления хромосом материнской звезды, при образовании второго имеющиеся хромосомы располагаются в две группы, и одна из них уходит вместе со вторым направляющим пузырьком. Таким образом, зрелое яйцо, как и готовый сперматозоид, содержит только половину хромосом, сравнительно с остальными телесными клетками, и сущность процесса оплодотворения сводится к восстановлению в яйце нормального для телесных клеток числа хромосом. Переходим теперь к процессу собственно оплодотворения. Тогда как у Physa весь сперматозоид проникает в яйцо, у большинства животных в яйцо проникает только головка и ближайшая к ней часть сперматозоида. Во всяком случае, после проникновения сперматозоида в яйцо его хвостик резорбируется, и в яйце, при конце его созревания или после созревания, видны только ядро и центросома сперматозоида. Последняя поворачивается, ложась между ядром яйца (женским) и ядром сперматозоида (мужским), и делится на две, развивая вокруг себя лучистость. Мужское ядро растет; женское передвигается от периферии, где было при образовании направляющих пузырьков, к центру яйца и здесь встречается с мужским ядром, после чего они сливаются, образуя ядро оплодотворенного яйца, тогда как центросома яйца еще ранее исчезает. Пройдя стадию клубка, слившиеся ядра образуют веретено с звездой хромосом, после чего начинается деление яйца, ничем не отличающееся от непрямого деления других клеток.
Деление яйца. Зародышевые листки. Оплодотворенное яйцо делится, т. е. распадается на т. н. шары дробления, или бластомеры, которые, делясь далее, становятся клеточками, дающими начало различным тканям и органам сложного организма. В зависимости от количества и способа распределения питательного желтка в теле яйца, деление происходит тем или иным способом, но питательный желток принимает лишь пассивное участие в процессе деления; активно делится только образовательный желток, или протоплазма тела яйца. Если питательного желтка немного, и он равномерно распределен в яйце, последнее все делится сначала на 2, потом на 4, 8 и т. д. бластомеров. Такое деление называется полным. Если же питательный желток преобладает количественно над образовательным, последний при делении не может увлечь его за собой, и потому в то время как количество бластомеров увеличивается, питательный желток почти не изменяется. Такое деление называется частичным или неполным. И тот и другой вид деления представляют многочисленные изменения, из которых надо упомянуть хотя бы тот случай, когда питательный желток занимает внутренние части яйца и образует — поверхностные; или — другой, когда протоплазма с ядром затеряна внутри питательного желтка. В первом случае деление происходит поверхностно, во втором — бластомеры появляются внутри яйца и уже оттуда перебираются на его поверхность. Так или иначе, делится яйцо, но в результате деления получается зародыш или только из бластомеров или из бластомеров, стоящих в известном отношении к питательному желтку. Смотря по способу расположения бластомеров, зародыш получает разные названия, как-то: morula, — если бластомеры представляют шаровидную кучку; blastula, — если зародыш походит на шар, с однослойной стенкой и полостью внутри; planula, — если слой призматических бластомеров (клеток) окружает внутреннее скопление округлых бластомеров; gastrula, — если зародыш имеет овальную форму и двухслойные стенки, окружающие полость, которая особым отверстием открывается наружу, тогда как внутренний слой стенок непосредственно переходит во внешний по краям этого отверстия. Стенка бластулы называется бластодермой; гаструла развивается из бластулы путем впячивания части стенки последней, после чего внешний слой стенок гаструлы называется эктодермой, а внутренний — энтодермой. Но кроме впячивания, гаструла может развиться последовательно из морулы и планулы путем расплывания большей части внутренней клеточной массы последней и прорыва в одном месте стенок такого двухслойного зародыша. Следовательно, схематично можно принять, что зародыш первоначально бывает однослойным (бластодерма), а позднее двухслойным (эктодерма и энтодерма), — и это может быть признано доказанным для зародышей всех животных. Слои, образующие стенки тела зародыша, называются зародышевыми листками. После того как экто- и энтодерма образовались, между ними, развивается мезодерма, составляющая третий зародышевый листок. Мезодерма происходит или на счет экто- и энтодермы, путем внедрения их клеток в промежуток между ними, или из мешковидных выпячиваний энтодермы. Если бластула есть полый шар, гаструла может быть сравнена с двустенным мешком, внешняя стенка которого (эктодерма) есть стенка тела и внутренняя — стенка кишечного канала, т. к. полость гаструлы является зачаточной пищеварительной полостью. Некоторые животные даже в развитом состоянии (например, гидроидные полипы) близки к гаструле по своему строению и, подобно ей, имеют только одно отверстие, ведущее в пищеварительную полость. Способ развития гаструлы из бластулы показывает, что полость бластулы соответствует промежутку между эктодермой и энтодермой, тогда как полость гаструлы есть новообразование. Следующей за гаструлой формой зародыша может быть принята трехслойная овальная пластинка, которая, заворачиваясь своими краями по продольной оси навстречу друг другу, дает три вставленные одна в другую трубки: внешнюю — из эктодермы, и внутреннюю — из энтодермы; что же касается мезодермы, то последняя, в свою очередь, расщепляется на внешний слой, который отходит к эктодерме, и внутренний — к энтодерме, тогда как между ними получается полость — т. н. полость тела. Только на спинной стороне зародыша, по направлению его продольной оси, мезодерма не расщепляется. Пищеварительная полость теперь открывается наружу двумя отверстиями: на переднем конце — ротовым и на заднем — заднепроходным. Из трех зародышевых листков развиваются все ткани и органы животного, а именно: из эктодермы - поверхностный слой общих покровов, связанные с кожей железы, существенные части органов чувств и центральная нервная система; из энтодермы — железистая выстилка кишечного канала и его придатки, а равно и некоторые части кровеносной системы; из мезодермы — все остальные органы, т. е. внутренний скелет, мышцы, большая часть кровеносной системы и т. д. Следовательно, при построении тела, например, позвоночного животного, наибольшее участие количественно принимает мезодерма. Она же, вместе с эктодермой, образует стенки тела (соматоплевра) и, вместе с энтодермой, стенки кишечного канала (спланхноплевра). Дальнейшее развитие зародыша из трубчатой формы состоит в постепенной дифференцировке трех зародышевых листков, причем рука об руку происходит дифференцировка тканей и органов. Установление учения о зародышевых листках, общих всему животному царству, является одним из наиболее значительных приобретений зоологии во второй половине XIX века.
Ткани. Как сказано, все высшие животные развиваются из единственной клетки — оплодотворенного яйца — путем последовательного деления. Первоначально происшедшие путем деления яйца клетки похожи друг на друга, имеют характерную для эмбриональных клеток приблизительно шаровидную форму и не различаются по отправлениям. Но по мере того как развитие продолжается, клетки все более и более отличаются друг от друга: они начинают дифференцироваться. Тогда как у простейших клетка выполняет все функции, у Metazoa, начиная с известной стадии, клетки утрачивают эту способность и дифференцируются на группы, из которых каждая способна выполнять только определенную функцию. Такая дифференцировка основана на принципе разделения труда. Группы клеток, дифференцированных в известном направлении, способных выполнять только определенные функции и связанных определенным способом, образуют ткани. Однако, ткани состоят не только из клеток, но и из межклеточного вещества, которое происходит из клеток, являясь то их выделением, то результатом превращения поверхностных частей протоплазмы клеток. В эмбриональном состоянии межклеточного вещества не образуется. Ткани делятся на пять категорий: 1) эпителиальную, 2) остовную, 3) мышечную, 4) нервную, 5) кровь и лимфу. Эпителиальная ткань состоит из плотно прилегающих друг к другу клеток, с ничтожным количеством связывающего их межклеточного вещества. Клетки обыкновенно лишены оболочки и имеют разную форму, благодаря чему устанавливаются разные виды эпителиальной ткани. Эпителий покрывает наружную поверхность тела и выстилает внутренние полости, являясь то плоским, то цилиндрическим, то однослойным, то многослойным. Если эпителий несет выделительную функцию, он называется железистым; если он воспринимает известные раздражения, которые передает нервной системе, то — чувствующим. У цилиндрического эпителия высота клеток более поперечника, у плоского — обратно. Цилиндрический эпителий, клетки которого несут на свободной поверхности реснички, называется реснитчатым. С железистым эпителием связаны железы двоякого рода: одноклеточные, или железистые, клетки и многоклеточные, имеющие вид трубчатых, мешковидных и гроздевидных желез. Железистая клетка одета оболочкой с отверстием, через которое выходит ее выделение. Последнее вырабатывается из плазмы, и только на дне такой клетки можно найти неизмененную плазму с ядром. Группа остовной ткани, или соединительных веществ, играет роль скелета в организме, а также служит для соединения разных частей тела или органов в одно целое, и в связи с такой своей функцией представляет весьма большое разнообразие. Отличительное свойство этих тканей составляет присутствие большого количества межклеточного, или основного, вещества, вследствие чего клеточные элементы ткани иногда играют совершенно второстепенную роль. Остовная ткань делится, в свою очередь, на: а) соединительную ткань, b) хрящевую ткань и с) костную ткань. Несмотря на большую разницу в их строении, они связываются друг с другом по происхождению. Все они закладываются в виде скопления эмбриональных клеток, округлой формы, без всякого признака промежуточного вещества, которое является несколько позднее в качестве продукта клеток. Соединительная ткань отличается присутствием межклеточного вещества, которое не бывает особенно твердым, и, в свою очередь, делится на несколько видов. Эмбриональная соединительная ткань, иначе называемая студенистой и слизистой, состоит из округлых или звездчатых клеток, соединенных между собой отростками, между которыми содержится в большом количестве гомогенное, содержащее слизь, промежуточное вещество. С возрастом животного, в основном веществе этой ткани появляются соединительно-тканные волоконца. Сетчатая соединительная ткань состоит исключительно из звездчатых клеток, анастомозирующих между собой отростками, благодаря чему получается сеть, ячеи которой заняты лимфой. Волокнистая соединительная ткань отличается строением своего промежуточного вещества, которое содержит соединительно-тканные и эластические волокна. Первые состоят из волоконец, склеенных в пучки или волокна особым цементом, и узнаваемы по своему отношению к реактивам. Эластические волокна встречаются гораздо реже первых, часто делятся дихотомически и легко узнаются уже по своей сильной светопреломляемости и эластичности. Клетки, в свою очередь, делятся на неподвижные, или прикрепленные к месту, и блуждающие. Первые имеют веретеновидную форму, но с возрастом животного становятся тоньше и тоньше. Некоторые из них содержат в себе зернышки красящего вещества (пигмента) и называются пигментными. Их форма обыкновенно звездчатая, и отростки, отходящие от тела клетки, способны к движению. Блуждающие клетки являются собственно лейкоцитами, или белыми кровяными тельцами, проникающими через стенки кровеносных сосудов в окружающие их ткани и передвигающимися в разных направлениях. Таким образом, блуждающие клетки могут быть встречены в разных тканях, но особенно многочисленны они в соединительной ткани. Та ткань, в основном веществе которой содержится большое количество эластических волокон, называется эластической. Иногда этих волокон бывает так много, что они совершенно закрывают собой соединительно-тканные, придавая всей ткани весьма своеобразный вид. Наконец, жировая ткань представляет собой такую соединительную ткань, клетки которой отлагают в себе более или менее значительное количество жира. Последний появляется в теле клетки в виде маленьких капелек, но потом, по мере увеличения их числа, они сливаются между собой, образуя капельки больших размеров, и, наконец, образуют одну большую каплю, которая оттесняет ядро клетки в ничтожные остатки протоплазмы тела. Хрящевая ткань отличается от соединительной большей твердостью межклеточного вещества, благодаря чему хрящ является переходом к костной ткани. Кроме того, для хряща является характерным строение и расположение хрящевых клеток и, наконец, образование при варке хондрина, или хрящевого клея. Самый обыкновенный вид хряща — т. н. гиалиновый, или стекловидный хрящ. Его основное вещество прозрачно, кажется однородным и значительно преобладает над клетками. Последние — округлой или овальной формы — часто бывают собраны в группы и иногда обнаруживают весьма правильное расположение. Тонкозернистая протоплазма содержит большое ядро с ясной оболочкой и ядрышками. У некоторых животных хрящевые клетки дают отростки, принимая звездчатую форму. Размножение обыкновенно происходит митотически, но иногда наблюдается и прямое деление. Хрящевые клетки лежат в основном веществе в ячеях, совершенно выполняя их собой. Та часть основного вещества, которая непосредственно прилежит к клетке, отличается своей большей светопреломляемостью, особенным сродством к красящим веществам и называется хрящевой капсулой. История развития хряща учит, что эмбриональные клетки сначала одеваются каждая оболочкой, потом эти оболочки утолщаются и сливаются, и таким образом из них получается основное вещество. По мере увеличения количества последнего, клетки все более и более расходятся между собой. Следовательно, хрящевые капсулы представляют собой оболочки клеток, еще не вполне слившиеся с уже однородным основным веществом, а их слоистость указывает на их происхождение и способ их роста. Иногда внутри капсулы видны две клетки, что указывает на деление клеток внутри капсулы. Снаружи хрящ одет слоем волокнистой соединительной ткани, т. н. надхрящницей, внутренней своей стороной непосредственно продолжающейся в хрящ. Рост хряща идет двумя способами: появлением новых слоев хряща со стороны надхрящницы (аппозиция) и увеличением количества основного вещества хряща изнутри, при участии хрящевых клеток (интусусцепция). В старости у млекопитающих основное вещество хряща может пропитываться известью, и тогда хрящ становится твердым, но ломким. У некоторых других животных хрящ в определенных местах нормально содержит известковые отложения (у акул). Если в основном веществе хряща содержатся ясно видимые соединительно-тканные волокна, он называется волокнистым; присутствие там же эластических волокон дает эластический хрящ. Костная ткань является самой твердой из остовных тканей и, за исключением зубной эмали, наиболее твердой ткани в организме позвоночных, почему играет роль опоры для мягких частей, образуя скелет животного. Подобно хрящу, костная ткань состоит из основного вещества и находящихся в нем клеток; но основное вещество, в свою очередь, состоит из органической части, т. н. костного хряща, и пропитывающих его углекислых и фосфорнокислых солей извести. Поэтому костная ткань может быть изучаема двояко: удалив действием слабой кислоты известь, получают органическое промежуточное вещество с заключенными в него костными клетками, которое, подобно хрящу, режется свободно ножом; или, высушив кость, при осторожном нагревании, сохраняют ее минеральные части, по удалении большей части органического вещества, после чего кость может быть изучаема на тонких шлифах. Таким образом, можно ознакомиться далее с тончайшим строением кости. На разных костях можно убедиться, что костная ткань может быть очень плотной (компактная костная ткань) и более рыхлой (губчатая костная ткань). Последняя напоминает собой скелет губки. И та и другая ткань обыкновенно бывают соединены в одной кости, но способ соединения их в разных костях различен. Изучая шлиф компактной костной ткани, сделанный в направлении длинной оси какой-либо трубчатой кости, например, фаланги пальца, при слабом увеличении, мы видим, что она пронизана вдоль каналами, которые соединяются друг с другом косыми анастомозами; это — т. н. Гаверсовы каналы, в которых проходят кровеносные сосуды. Система этих каналов, с одной стороны, открывается в определенных местах наружу, с другой — в большую полость внутри кости. В основном веществе находятся во множестве микроскопически малые полости, костные тельца, занятые костными клетками. На поперечном шлифе через ту же кость Гаверсовы каналы в поперечном разрезе являются правильно округлыми, и только соединяющие их ветви — овально-вытянутыми. Тогда как на продольном шлифе костные клетки расположены рядами, более или менее параллельно Гаверсовым каналам, на поперечном шлифе они образуют по отношению к последним концентрические ряды. Основное вещество кости имеет пластинчатое строение и состоит из нескольких систем пластинок, очерчивающих Гаверсовы каналы, параллельных наружной и внутренней поверхности кости и выполняющих промежутки между теми и другими. Поверхность кости одета волокнистой соединительной тканью, надкостницей (periostium), которая продолжается в систему гаверсовых каналов, выстилая их стенки, и доходит до внутренней полости кости. Последняя полость занята особой жировой тканью, т. н. костным мозгом, почему называется костномозговой полостью. Полости, содержащие костные клетки, дают от себя отростки, которыми не только соединяются между собой, но также с поверхностью кости, Гаверсовыми каналами и костномозговой полостью. Таким образом, компактная костная ткань пронизана множеством полостей разного размера, содержащих как кровеносные сосуды, так и клетки, и этим обусловливается питание костной ткани и обмен веществ. Особенно легко убедиться в таком строении костной ткани, наполнив описанную систему полостей каким-либо покрашенным веществом. Костные клетки лишены оболочки и по форме точно соответствуют содержащим их полостям, совершенно выполняя их. Следовательно, они несколько вытянуты в направлении продольной оси кости. Давая от себя отростки в канальцах, соединяющих костные тельца, костные клетки приобретают звездчатую форму. Эти отростки клеток соединяются друг с другом у одних животных в течение всей жизни, у других — только в молодости. Тончайшее строение губчатой костной ткани существенно не отличается от строения компактной костной ткани, но в губчатой ткани нет Гаверсовых каналов и нет правильных систем пластинок основного вещества, хотя пластинчатость наблюдается в более толстых балках губчатой ткани. Мышечная ткань отличается способностью своих элементов сокращаться лишь в одном направлении. Это сокращение может происходить как под влиянием воли животного, так и без этого фактора, и потому физиологически мышцы могут быть разделены на подчиненные волевым импульсам и им не подчиненные. Гистологически первые соответствуют поперечнополосатым мышцам, вторые — гладким. Особое место занимает сердечная мышца, по строению примыкающая к первым, но не подчиненная волевым импульсам. Гладкие мышцы состоят из веретенообразных клеток, лишенных собственной оболочки. В средней утолщенной части клетки лежит овальное или палочкообразное ядро, с одним или несколькими ядрышками, а по концам его собрана зернистая протоплазма. В остальной части тела клетки видны идущие по длинной оси последней сократимые волоконца, находящиеся в недифференцированной саркоплазме. Гладкие мышечные волокна некоторых органов способны временно достигать очень больших размеров (гипертрофироваться) и потом опять возвращаться к прежней величине (матка). Описываемые элементы иногда лежат отдельно, иногда образуют подобие оболочек, благодаря своему расположению слоями, причем, то лежат рядом друг с другом, в одном направлении, то перекрещиваются. Отдельные волокна таких скоплений склеены между собой особым цементом. Сокращение гладких мышечных волокон происходит медленно. Они находятся в стенках кишечного канала, в органах дыхания, в мочеполовом аппарате, в сосудистой системе, в некоторых железах, в коже и глазе. Поперечнополосатые мышечные волокна представляют собой весьма высоко дифференцированный вид ткани. Образующие их многоядерные клетки вытянуты в длинные волокна с поперечной и продольной полосатостью, из которых наиболее характерна и резко выражена первая. Тогда как гладкие мышечные волокна имеют длину от 50 до 200 μ, при толщине в 4—7 μ, и только гипертрофированные волокна бывают до 500 μ, поперечнополосатые мышечные волокна, при сравнительно небольшой толщине, в 30—60 μ, могут достигать длины свыше 10 см. Из поперечнополосатых мышечных волокон образованы мышечные пучки, или мышцы, одевающие собой внутренний скелет позвоночных животных и находящиеся внутри от наружного скелета в теле членистоногих. Поперечнополосатые мышечные волокна могут оканчиваться или тупо-округло или заостряясь конусообразно. Иногда концы волокна бывают расщеплены. Поперечная и продольная полосатость обусловливается внутренним строением волокна, которое состоит из продольных волоконец, каждое с поперечной полосатостью. Волоконца идут параллельно друг другу от одного конца волокна до другого, сократимы, — в зависимости от чего сократимо все волокно, — но связаны остатками неизмененной протоплазмы (саркоплазма). Волоконца расположены в волокне или всюду равномерно или пучками. В последнем случае на поперечном разрезе волокна видны т. н. поля Конгейма, состоящие из участков, занятых пучками перерезанных волоконец и находящихся между ними прослоек саркоплазмы. У низших животных саркоплазмы больше, нежели у высших. В саркоплазме находятся овальные ядра, своей продольной осью лежащие по длине волокна. В одних волокнах ядра лежат в периферической саркоплазме, в других — между пучками волоконец, что зависит от количества саркоплазмы. Так, у человека наблюдается первый случай, у лягушки — второй. У некоторых животных имеются т. н. белые и красные мышцы, различающиеся также количеством саркоплазмы и положением ядер: у первых саркоплазмы мало, и ядра лежат на периферии; у вторых ее много, и ядра лежат внутри волокна. Красные мышцы медленнее сокращаются и позднее утомляются. У большинства позвоночных почти исключительно имеются мышцы, соответствующие красным. Каждое мышечное волокно одето очень тонкой, бесструктурной оболочкой — сарколеммой, которая соответствует оболочке клетки и облекает волокна со всех сторон, не исключая его обоих концов. Только в виде исключения у некоторых низших животных нет сарколеммы. Поперечная полосатость мышечных волокон обусловливается поперечной волосатостью волоконец, при условии расположения физически одинаковых отрезков последних на одном и том же уровне. Физическое различие в строении отрезков внешним образом выражается в том, что одни из них сильнее преломляют свет, другие — слабее. Первые красятся интенсивнее, нежели вторые, и при низкой установке трубки микроскопа кажутся более темными, нежели промежутки между ними. Темные отрезки, или поперечные диски, состоят из оптически анизотропного вещества, т. е. обладают способностью двойного светопреломления; вторые (светлые) — из оптически изотропного вещества, т. е. с простым светопреломлением. Детальное изучение поперечнополосатых мышечных волокон показывает, однако, гораздо более сложную структуру, наилучше изученную, между прочим, на мышцах майского жука. Особенно легко убедиться в сложном строении поперечных дисков при одностороннем сокращении мышечного волокна. Наблюдения показали, что при сокращении мышцы поперечные диски являются деятельными, уменьшаясь в объеме и уплотняясь вследствие отдачи жидкости окружающему веществу. Однако, укорачиваясь, они становятся в то же время несколько шире. С этим совпадает и то обстоятельство, что целая мышца, состоящая из большего или меньшего количества мышечных волокон, при сокращении укорачивается и утолщается. Изотропное вещество при сокращении, будучи эластично, играет лишь пассивную роль. Поперечная полосатость не стоит в качестве необходимого условия сокращения, т. к. гладкие мышечные волокна также сокращаются, но имеет прямое отношение к быстроте сокращения. Саркоплазма, не участвуя прямо в деятельности мышечного волокна, имеет значение для его питания, размножения и роста. Она же является проводником нервного раздражения и его равномерной передачи всем частям волоконец. Клетки, из которых развиваются поперечнополосатые мышечные волокна, сначала одноядерны и имеют веретенообразную форму, но затем клетка удлиняется, при одновременном делении ядра, и принимает вид волокна, в котором еще позднее появляется характерная структура. Мышца, в зависимости от своей величины, может состоять из большего или меньшего количества мышечных волокон, связанных вместе соединительной тканью. Небольшие партии мышечных волокон носят название мышечных пучков, а последние, в свою очередь, складываются в мышцу, которая снаружи также одета соединительнотканной оболочкой. Мышечная ткань легко разрушается и легко восстановляется. Восстановление происходит путем расщепления уже готовых мышечных волокон, причем мышечные ядра начинают энергично размножаться и образуют несколько продольных скоплений, по числу которых одно волокно распадается на несколько новых. Нервная ткань состоит из клеток и волокон, из которых последние являются отростками первых. Каждая нервная клетка с ее отростками называется нейроном, и отростки одной нейронной клетки могут только соприкасаться с отростками другой, отнюдь не переходя в них непосредственно. В зависимости от числа отходящих от нее отростков, нейронная клетка может быть грушевидной, веретеновидной или звездчатой. По числу отростков нейронные клетки различаются как униполярные (с одним), биполярные (с двумя) и мультиполярные (со многими). По строению отростки нейронных клеток делятся на две категории: нервные отростки, иначе нейриты, дейтерсовы отростки, или осевые цилиндры, — имеют вид нежных, тонких, с гладким краем, волокон, которые могут достигать значительной длины (иногда свыше 1 м); протоплазматические отростки, или дендриты, — имеют неровный край и уже на коротком расстоянии обыкновенно дают обильное ветвление. Концевые разветвления тех и других отростков называются телодендриями. Каждая нейронная клетка имеет один нервный отросток; следовательно, униполярные клетки — только с нейритом, биполярные — с одним нейритом и одним дендритом, мультиполярные — с одним нейритом и несколькими дендритами. Нейриты имеют значение проводящих путей нейрона; дендриты рассматриваются в качестве его органов питания. Тело нейронной клетки, независимо от формы, имеет фибриллярное строение, причем фибрилли продолжаются в него из отростков, располагаются концентрически и, давая от себя отростки, часто образуют около ядра сеть. Тело нейронной клетки содержит ядро очень характерной формы: большое, пузыревидное, с ясно выраженной оболочкой. Обыкновенно в нем видно также ядрышко. Центросома найдена лишь в некоторых нейронных клетках. Собственно оболочки нейронная клетка не имеет, но периферически лежащие нейронные клетки часто получают вторичные оболочки соединительнотканного происхождения. Нервное волокно состоит из осевого цилиндра, т. е. удлинения отростка нейронной клетки, и одевающих его оболочек. Осевой цилиндр идет непрерывно от места его отхождения от нейронной клетки до конца волокна, имеет фибриллярное строение и содержит между волоконцами ничтожное количество неизмененной плазмы (нейроплазма). Он очень нежен и чрезвычайно восприимчив к реактивам. Оболочки нейронных волокон хорошо видны на поперечном разрезе через последнее и состоят из мозжечкового влагалища, непосредственно прилегающего к осевому цилиндру, и Шванновской оболочки, облекающей первое. Мозжечковое влагалище состоит из миэлина, жироподобного, вязкого, сильно преломляющего свет вещества, которое образует вокруг осевого цилиндра толстую оболочку, почему последняя в оптическом разрезе нейронных волокон является его двухконтурной оболочкой. Эта оболочка прерывается через определенные промежутки, чем обусловливается существование т. н. перетяжек Ранвье, где Шванновская оболочка непосредственно подходит к осевому цилиндру, а в промежутках между ними мозжечковое влагалище косо прорезано как бы насечками (Шмидт-Лантермановские насечки), которые дают подобие конических отрезков, вставленных друг в друга. По обработке вытягивающим жир реактивами, миэлин является в виде сети, облекающей осевой цилиндр, с ясными перерывами в области перетяжек Ранвье. Эта сеть состоит из вещества, напоминающего рог и называемого нейрокератином. Шванновская оболочка имеет вид тонкой перепонки с ядрами, около которых видно некоторое скопление протоплазмы. Нервные волокна ветвятся, причем всегда дают ветви в перетяжках Ранвье. Кнаружи от Шванновской оболочки может лежать соединительнотканная оболочка или оболочка Генле с рассеянными в ней ядрами. Мозжечковое влагалище, вероятно, играет роль изолятора для нервных волокон, так как чувствительность последнего увеличивается с возрастом, по мере развития мозжечкового влагалища. Некоторые нервные волокна одеты только мозжечковым влагалищем, другие только Шванновской оболочкой. Нервные клетки развиваются из округлых эмбриональных клеток (нейробласты), между которыми нет промежуточного вещества. Одни из этих клеток развиваются в нервные клетки, другие — в связывающие последние клетки особой ткани, нейроглии, имеющей большое значение при построении центральной нервной системы. Последний вид тканей составляют кровь и лимфа, которые можно рассматривать как ткань с жидким межклеточным веществом. Кровь высших животных представляется жидкостью красного цвета, которая состоит из кровяной плазмы (межклеточное вещество; и форменных элементов: кровяных телец, кровяных пластинок и первичных зернышек. Между кровяными тельцами различаются красные и белые. Красные кровяные тельца, или эритроциты, содержат гемоглобин, благодаря которому кровь в тонких слоях окрашивается в желтоватый цвет, часто с зеленоватым оттенком, а в более толстых в характерный красный цвет. У млекопитающих красные кровяные тельца в огромном большинстве случаев — сплющенные, округлые тельца, вогнутые с обеих сторон, отчего в профиль кажутся бисквитообразными. Только в семействе верблюдов они — овальные, как у остальных позвоночных, но у верблюдов, как и у остальных млекопитающих, они без ядер, тогда как у птиц, рептилий, амфибий и рыб содержат овальные ядра (каждое — одно). Отсутствие ядер одними объясняется как результат механического процесса — прямым выхождением ядра из эритроцита, другими — как результат превращения ядра внутри последнего. Во всяком случае, первоначально, на ранних стадиях, зародыш млекопитающих содержит ядерные эритроциты, которые только потом становятся безъядерными. Красные кровяные тельца лишены оболочки и состоят из протоплазматического основного вещества, т. н. стромы, и красящего вещества, гемоглобина, характеризующегося большим сродством к кислороду. Следовательно, красные кровяные тельца являются настоящими клетками, в одном случае — ядерными, в другом — безъядерными. Они очень чувствительны к внешним раздражениям, растяжимы, эластичны и обладают способностью в крови, взятой из кровеносного сосуда, складываться своими плоскими сторонами, образуя подобие монетных столбиков. Количество их в 1 куб. мм очень различно у разных животных, варьируя от 33 000 (у протея) до 19 000 000 (у козы). У человека в 1 куб. мм около 5 000 000 эритроцитов, но это количество подвержено колебаниям даже в нормальных условиях. Наибольшее их количество наблюдается сейчас же после рождения, а затем начинает уменьшаться. Белые кровяные тельца, или лейкоциты, лишены оболочки, содержат одно или несколько ядер и лишены определенной формы, т. к. способны совершать амёбообразные движения. В состоянии покоя они округлы. Количество их в крови менее постоянно, нежели эритроцитов, и потому не может быть выражено в приблизительно точных цифрах. Лейкоциты могут выходить из кровеносных сосудов, проходя через их стенки между клетками, и таким образом дают начало т. н. блуждающим клеткам. Первичные зернышки обладают всеми свойствами эритроцитов и как бы являются ими в миниатюре. Кровяные пластинки представляют собой очень нежные, бесцветные, блестящие, округлые или овальные пластинки, которые, по мнению большинства, играют большую роль при свертывании крови. Кровяные тельца развиваются вместе с появлением кровеносных сосудов. Развиваются ли белые и красные кровяные тельца из одной основной формы клеток или из разных, остается нерешенным. Красные кровяные тельца развиваются из эритробластов — округлых ядерных клеток, несколько крупнее безъядерных эритроцитов. Делясь повторно, они дают начало ядерным красным кровяным тельцам, а затем из последних получаются безъядерные. Развитие красных кровяных телец в течение эмбриональной жизни происходит в печени, в стенках пупочного канатика, в красном костном мозге, в селезенке и отчасти, вообще, в лимфатических железах. Особенно важна в этом отношении печень. У взрослых образование эритроцитов происходит почти исключительно в красном костном мозге. Лейкоциты появляются в крови позднее эритроцитов, вместе с развитием лимфатических желез. Кровь свертывается благодаря присутствию в ее плазме особого белкового вещества — фибрина, которое при свертывании крови распадается на две составные части. Лимфа отличается от крови своей бесцветностью и состоит из прозрачной жидкой плазмы лимфы и содержащихся в ней белых телец — лимфоцитов, не отличающихся от лейкоцитов. Других клеточных элементов в ней нет.
Органы. Тело сложных животных состоит из разных частей, или органов, несущих определенные отправления. С этой точки зрения и ткани можно считать за простейший вид органов, но обыкновенно они рассматриваются сами по себе и, вместе с тем, как составные части органов в собственном смысле слова. Последние делятся на следующие группы: 1) общие покровы, 2) органы движения, 3) органы пищеварения, 4) органы дыхания, 5) органы кровообращения, 6) органы выделения, 7) органы размножения, 8) нервная система и 9) органы чувств.
1) Общие покровы одевают тело животных снаружи и защищают от вредного воздействия внешней среды. В простейшем виде они представлены только эпителиальным слоем, клетки которого расположены в один ряд и покрыты на их свободной поверхности кутикулой или, напротив, мерцательными ресничками. Иногда кутикула заменена хитином, который может быть очень толст и периодически сбрасывается в своих поверхностных частях (линька). Иногда хитин соединяется с известковыми солями, и тогда тело животного оказывается заключенным в панцирь, гнущийся только в определенных частях, где покровы остаются мягкими. Своеобразная мантия оболочников иногда по консистенции и структуре напоминает хрящ, но ее вещество развивается как выделение эктодермических клеток, часть которых позднее находит себе в ней место. Затем общие покровы осложняются прибавлением к их эпителиальному слою подстилающего его соединительнотканного, а также выделением сложного наружного скелета из органической основы с известковыми солями. Таковы, например, общие покровы моллюсков, выделяющие раковину, которая состоит из трех слоев. Среди обыкновенных эпителиальных клеток покровов, у многих животных встречаются железистые клетки. Однослойный эпителий характеризует собой покровы беспозвоночных и у позвоночных во взрослом состоянии известен только у ланцетника. У всех остальных позвоночных общие покровы состоят из многослойного эпителия, или эпидермиса, и у всех, не исключая ланцетника, из соединительнотканного слоя кожи собственно. Эпителиальный слой построен различно у водных и наземных позвоночных. У водных он несет на поверхности кутикулу или совершенно свободен, у наземных кутикулу заменяет поверхностный слой рогового эпителия. Эпидермис водных позвоночных содержит железистые клетки, эпидермис наземных дает начало сложным железам — мешковидным, трубчатым и гроздевидным, которые, не умещаясь в нем, проникают в более глубокие слои. При дифференцировке поверхностного эпидермиса в роговой, его глубокие слои называются слизистым или Мальпигиевым эпидермисом. Глубокие клетки последнего способны к размножению, тогда как поверхностные, в виде роговых чешуй, постепенно отмирают и спадают. Соединительнотканный слой кожи может быть простым, или его глубокий слой дифференцируется в подкожную клетчатку, обыкновенно содержащую скопления жира. В слое кожи волокна иногда идут замечательно правильно, располагаясь в три системы перекрещивающихся балок, иногда же расположены совершенно неправильно. Твердые части общих покровов позвоночных могут принадлежать и эпидермису, и коже собственно; иногда те и другие существуют одновременно. У рыб эти твердые образования представлены чешуями, развивающимися в виде костных пластинок в слое собственно кожи, но у одной группы имеют вид костных пластинок, каждая с шипом, который одет на поверхности слоем эмали. У рептилий на поверхности тела развиваются роговые чешуи и щитки исключительно эпидермического происхождения, иногда подстилаемые костными образованиями кожи. У черепах те и другие образования достигают наибольшего развития, складываясь в два щита, спинной и брюшной, которые защищают тело соответственно сверху и снизу. У птиц роговые щитки и чешуйки сохраняются лишь местами, тело же одето перьями очень сложного строения, но развивающимися из зачатка, соответствующего роговой чешуе рептилий. У млекопитающих характерным эпидермическим образованием является волос, который, как и его видоизменения — щетина, иглы, развивается точно также из зачатка, соответствующего роговой чешуе. Местами у разных млекопитающих сохраняются и настоящие роговые щитки и чешуи, а у панголина тело, не исключая хвоста и конечностей, одето роговыми чешуями, из которых каждая является в результате срастания групп волосовидных образований.
2) Органы движения чрезвычайно разнообразны. С одной стороны, у водных животных низших групп они представлены реснитчатым покровом, или сплошным или локализованным на определенных участках. С другой, — выражены выростами и придатками тела, к числу которых относятся и конечности позвоночных. Наконец, у иглокожих органы движения представлены особой амбулякральной системой, которая состоит из полых трубочек и связанных с ними цилиндрических придатков, с присосками т. н. амбулякральных ножек. Эти трубочки могут то наполняться водой извне, то освобождаться от нее, и таким способом амбулякральные ножки приводятся в действие. Из реснитчатых органов движения особенно замечательны весельные пластинки ребровиков. Все выросты и придатки снабжены сильной мускулатурой, которая является необходимой принадлежностью органов движения большинства беспозвоночных и всех позвоночных. В том случае, если придатков, несущих функцию органов движения, нет, движение производится волнообразными изгибами всего тела, что достигается сокращением туловищных мышц. Земляные черви движутся, вытягивая и последовательно сокращая свое тело, при условии укрепления его переднего конца во время сокращения; при этом дополнительными органами движения служат ряды щетинок, торчащих из кожи. Моллюски движутся или при помощи выроста брюшной стороны тела — т. н. ноги, или при помощи особого органа — т. н. воронки, через которую выбрасывается вода, чем дается толчок телу животного, передвигающемуся в противоположном направлении. Так же прыжками, при соответствующем сокращении колокола, движутся медузы. Собственно конечности выражены у членистоногих и позвоночных и могут быть сведены к трем основным типам: водным, наземным и воздушным. Водные представлены весельными ножками ракообразных, плавниками рыб, ластами сирен и китообразных. Они всегда плоски, веслообразны, но могут очень варьировать в подробностях своей формы. Наземные конечности служат или для передвижения по земле, или для рытья в земле. Первые более или менее тонки и, если служат для прыгания, то длинны; таковы — задние ножки кузнечиков, задние конечности кенгуру, тушканчиков и пр. Роющие всегда широки, несколько плоски и массивны; таковы — передние ножки медведки, лапы крота и др. Воздушными органами движения служат крылья и особого рода парашюты. У насекомых крылья являются парными спинно-боковыми выростами 2-го и 3-го сегментов груди, но задняя пара может и недоразвиваться. В случае, если существуют обе пары, они могут быть одинаковы или различны. В последнем случае передняя пара — толстая, твердая, сильно хитинизированная, служит покрышкой для задней и называется элитрами. Задняя — является только органами летания и прозрачна. У позвоночных органы воздушного движения представлены тоже крыльями, развивающимися в результате видоизменения передних конечностей. У птиц это достигается изменением скелета конечностей и своеобразным расположением принадлежащих ей перьев, у млекопитающих — тоже изменением скелета конечностей и развитием между его частями особых перепонок кожного происхождения. Наконец, парашюты служат не для летания, а для прыгания с дерева на дерево на большое расстояние и представляют собой складки кожи между конечностями и туловищем, а также кпереди от передних и кзади от задних конечностей. Такие парашюты развиты у белок, у летающего маки.
3) Органы пищеварения в простейшем их виде не отделены от общей полости тела, что мы встречаем у кишечнополостных. У низших червей они представлены полостью разной формы, но имеющей лишь одно входное отверстие. Наконец, у высших червей, огромного большинства беспозвоночных и у позвоночных органы пищеварения представлены кишечным каналом различной длины, открывающимся спереди ротовым отверстием, сзади заднепроходным и расчлененным на ряд отделов, выполняющих специальные функции. Наибольшей сложности кишечный канал достигает у насекомых и млекопитающих. В связи с кишечным каналом стоят разные железы, принадлежащие к его определенным отделам, а именно слюнные, печень и поджелудочная (панкреас). У многих членистоногих в кишечный канал открываются Мальпигиевы трубочки, несущие функцию почек. В схеме подразделения кишечного канала у позвоночных представляются в следующем виде: за ротовым отверстием лежит полость рта, которой принадлежит челюстной аппарат с зубами, служащими для измельчения пищи. Ротовое отверстие ограничено сверху и снизу соответственно верхней и нижней губой, а слизистая оболочка рта иногда образует по бокам ротового отверстия защечные мешки, куда набивается корм, который животное поедает позднее или переносит в свои складочные амбары. У некоторых китов зубы существуют лишь в зародышевом состоянии, а потом исчезают, и их место занимает т. н. китовый ус. На дне ротовой полости сидит язык в виде мясистого придатка, а ее крышу составляет нёбо. За ротовой полостью следует глотка, обыкновенно довольно резко отграниченная от первой, а за последней идет пищевод, в виде трубки сравнительно простого устройства, но разной длины и ширины. Пищевод открывается в желудок, чрезвычайно разнообразного устройства, начиная от простого и кончая сложным желудком жвачных. Передний конец желудка называется кардиальным, задний — пилорическим. В случае сложного желудка, пища не прямо проходит от его переднего конца до заднего, а сначала наполняет два первые отдела, затем возвращается в рот и, будучи здесь пережевана и обильно смочена слюной, снова спускается в желудок, который и проходит теперь до его заднего конца. Последний имеет кольцевидный пилорический клапан. Начинающиеся за желудком кишки имеют чрезвычайно различную длину, будучи сравнительно короткими у хищных, и очень длинными у травоядных. Они делятся на тонкие, толстые и прямую. На границе между тонкими и толстыми находится слепая кишка. Прямая кишка открывается наружу у огромного большинства млекопитающих заднепроходным отверстием, но у низших млекопитающих, у птиц, рептилий, амфибий и некоторых рыб она впадает в клоаку, куда открываются, кроме того, протоки мочеполового аппарата. С ротовой полостью связаны подчелюстные, подъязычные и околоушные железы (слюнные). Печень, как и поджелудочная железа, открывается своими протоками (иногда вместе) в первую петлю тонких кишок, т. е. за желудком. Печень представляет собой массивную лопастную железу, часто с желчным пузырем, который играет роль резервуара, где собирается желчь, выделяемая печенью. Протоки печени называются общим именем желчных или, точнее, печеночных, проток желчного пузыря — пузырным, а проток, образующийся из соединения желчного с пузырным, — общим желчным протоком. Отношение протоков печени и желчного пузыря друг к другу и к кишечнику представляет несколько типов. Поджелудочная железа — нежная гроздевидная железа, лежащая в складке брыжейки и довольно трудно уловимая. У некоторых рыб кишка не образует петель и коротка, отчего ее поверхность усвоения была бы мало развита, если бы не было особого приспособления. Последнее выражено в т. н. спиральном клапане, т. е. в складке слизистой оболочки кишки, образующей спиральные извивы, благодаря которым пища продвигается через кишку достаточно медленно, чтобы вместе с тем быть всосанной пищеварительной поверхностью. У паразитных форм кишечный канал часто атрофируется более или менее полно, и питание происходит всасыванием питательной жидкости стенками тела.
4) Органы дыхания чрезвычайно разнообразны, то являясь внешними придатками, то системой внутренних полостей, иногда гроздевидных, иногда трубчатых. В простейшем виде дыхание совершается всей поверхностью тела, то есть в качестве органов дыхания функционирует кожа, но развитие на последней кутикулы, хитина и других твердых образований затрудняет ее дыхательную функцию, которая, таким образом, приурочивается только к определенным участкам. Смотря по тому, живет ли животное в воде или на суше, т. е. непосредственно в воздушной среде, мы имеем два основных типа органов дыхания: органы водного дыхания, каковыми являются жабры, и органы воздушного дыхания, куда относятся трахеи, легочные мешки и легкие. У некоторых животных существуют одновременно органы воздушного дыхания двоякого рода, например, легочные мешки и трахеи, и у немногих, живущих и в водной и в наземной среде, одновременно существуют даже органы водного и воздушного дыхания. Примером животных первой категории являются паукообразные, второй — амфибии. Жабры делятся на наружные и внутренние. Первые являются кожными выростами различной формы, с очень тонкими стенками и хорошо развитыми кровеносными сосудами внутри. Они могут быть нитевидны, листовидны, ветвисты или гребенчаты и принадлежать то разным областям тела, например, разным сегментам тела у кольчатых, то быть собранными на более ограниченных местах, например, переднем конце тела у тех же животных. У ракообразных жабры принадлежат некоторым членистым придаткам головогруди. У моллюсков жабры в основе нитевидны, но путем вторичных осложнений могут повести к образованию сложных ситовидных пластинок. У земноводных наружные жабры развиваются как ветвистые выросты по бокам шейной области над внутренними жабрами. Внутренние жабры развиваются у позвоночных в связи с передним концом кишечного канала, который в области глотки пронизан жаберными щелями, ведущими из глотки наружу. Жаберные лепестки сидят по краям этих щелей, образуя гребенчатые придатки. Жаберные щели могут быть или видимы снаружи или с каждой стороны прикрыты жаберной крышкой, развивающейся из складки кожи. Вода, входя через рот, проходит в глотку и отсюда выходит наружу, омывая жаберные щели. Трахеи представляют собой трубки своеобразного строения, начинающиеся попарно в каждом сегменте двумя боковыми отверстиями (стигмы) и обильно ветвящиеся внутри тела животного, где они проникают буквально к каждому органу, принося с собой запас воздуха. Количество стигм может уменьшаться вследствие атрофии некоторых из них, но расположение трахей существенно не изменяется, сводясь в типе к двум боковым продольным стволам и их боковым ветвям. Легочные мешки являются парными впячиваниями стенок тела в виде мешков, или карманов, дно которых образует многочисленные тонкие вторичные складки. Таким образом, получается подобие книги с несколько разошедшимися страницами. Воздух входит через отверстие в полость мешка и проникает снаружи между его складками, внутри которых содержится кровь, попадающая сюда из общей полости тела. Под легкими у разных животных разумеются существенно различные органы. У моллюсков это — губчатая ткань на определенном месте стенок мантийной полости. У позвоночных это — парный мешковидный орган, развивающийся со дна глотки, с которой связан особой дыхательной трубкой, или трахеей. Трахея на своем нижнем конце делится на две бронхи, а последние распадаются на более мелкие веточки, которые оканчиваются в дольках легкого пузырчатого строения. Воздух поступает в легкие и выводится отсюда через дыхательное горло, окисляя кровь, приносимую особыми сосудами в стенки легочных пузырьков. В связи с дыхательным горлом развивается голосовой аппарат. У птиц позади легких лежат воздушные мешки, соединенные с ветвями бронх и заходящие своими отростками в полости трубчатых костей, которые благодаря этому теряют свой костный мозг.
5) Органы кровообращения стоят в тесной связи с органами дыхания. Там, где нет особых органов дыхания, кровь выполняет собой общую полость тела или промежутки паренхиматозной ткани и здесь передвигается в разных направлениях, благодаря движению и сокращениям тела, без участия особого специального органа. Но в огромном большинстве случаев существует особый орган — сердце, которое своими сокращениями проталкивает кровь в определенном направлении. В простейшем виде сердце имеет вид мешка с передним, задним и боковыми отверстиями. Последние защищены клапанами, открывающимися внутрь. При сокращении такого сердца кровь выходит через его переднее и заднее отверстия, а при расширении входит в него через боковые отверстия. Снаружи сердце одето околосердечной сумкой (перикардий). При дальнейшем осложнении сердца края его переднего и заднего отверстий вытягиваются в трубки — выносящие сосуды, или артерии. Еще дальше вытягиваются в приносящие сосуды, или вены, края боковых отверстий сердца. На этих стадиях развития находим сердце у разных ракообразных. У насекомых оно удлиняется в трубку с несколькими парами боковых отверстий. Однако, среди беспозвоночных самая сложная система органов кровообращения имеется у моллюсков, где не только сердце продолжается в артерии и вены, но где вены в месте впадения в сердце расширяются, давая т. н. предсердия. Кроме того, артерии и вены распадаются на мелкие веточки, но, как ни близко артериальные веточки подходят к венозным, все-таки между ними остаются каналы без собственных стенок, где кровь проходит т. н. системой лакун. Такая кровеносная система, где тончайшие артериальные веточки не переходят непосредственно в венозные, и где кровь проходит часть своего пути лакунами, называется незамкнутой. Она принадлежит всем беспозвоночным. Напротив, у позвоночных кровеносная система — замкнутая, т. к. тончайшие разветвления артерий (артериальные капилляры) непосредственно переходят в тончайшие разветвления вен (венозные капилляры). У ланцетника сердца нет, и оно заменяется сократимостью крупных сосудов. У остальных позвоночных сердце есть и подводится к трем типам: сердце двухкамерное — с одним предсердием и одним желудочком, позади предсердия венозная пазуха, впереди желудочка иногда артериальный конус; сердце лежит позади органов дыхания, следовательно, через него проходит исключительно венозная кровь; сердце трехкамерное — с двумя предсердиями и одним желудочком, при венозной пазухе и, иногда, остатках артериального конуса; в левое предсердие поступает кровь из органов дыхания, следовательно артериальная, в правое из других органов, следовательно венозная; в желудочке оба потока крови отчасти смешиваются, но к головному мозгу идет преимущественно артериальная, к органам дыхания преимущественно венозная и только к остальным органам смешанная; сердце четырехкамерное — с двумя предсердиями и двумя желудочками, без каких бы то ни было внешних придатков; левая половина сердца (предсердие и желудочек) артериальная, правая (предсердие и желудочек) венозная; от левого желудочка кровь идет к правому предсердию по большому кругу кровообращения, омывая все органы, кроме органов дыхания, поступает далее, не изменяя своего состава, в правый желудочек и отсюда по малому кругу кровообращения, пройдя через органы дыхания, подходит к левому предсердию; таким образом, устанавливается деление четырехкамерного сердца на правую — венозную и левую — артериальную половину. У позвоночных наряду с кровеносной системой существует лимфатическая, состоящая из ряда сосудов, которые начинаются млечными ходами в стенках кишок и оканчиваются, впадая в яремные вены. Лимфатическая система существенно дополняет собой кровеносную в деле усвоения организмом пищевых веществ.
6) Органы выделения могут отсутствовать в качестве особой системы, что обыкновенно стоит в связи с отсутствием в то же время обособленных органов дыхания и кровообращения. Однако, это не значит, что там, где нет органов выделения, нет и выделительной функции: подобно тому как дыхание и питание возможны без специальных органов, и выделение может происходить без т. н. органов выделения. Там, где они есть, их функция сводится к удалению из организма ненужных веществ в жидком виде. Органы выделения всех животных могут быть сведены к трем формам, из которых две, несмотря на кажущееся различие между ними, могут быть сведены к одному типу. В простейшем виде органы выделения представлены бесчисленными ветвящимися канальцами, которые все сходятся в один непарный или два парных канала, сравнительно значительного диаметра, открывающихся на заднем конце тела. Каждая веточка этой системы канальцев оканчивается воронкообразным расширением, которое или находится свободно в общей полости тела или погружено в занимающую место общую полость тела паренхиматозную ткань. Каждая воронка прикрыта особой клеткой, от которой внутрь воронки спускается большой жгутик. Подобные же жгутики наблюдаются местами и на остальном протяжении выделительных каналов, способствуя своими движениями проведению жидкости в определенном направлении. Клетки на воронках, несомненно, относятся к категории железистых клеток, и не просто фильтруют жидкие вещества, выводимые из организма, а выделяют их в буквальном смысле слова. Органы выделения второго типа представлены т. н. сегментальными органами, или нефридиями, которые являются посегментно расположенными парами трубочек, одним концом связанных со стенками тела, где они открываются наружу боковым отверстием, а другим — свободно оканчивающихся в полости тела. На своем пути сегментальный орган проходит через перегородку между двумя сегментами, которая укрепляет его; своим передним концом он принадлежит одному сегменту, задним — другому, сзади лежащему. Следовательно, в каждом сегменте в действительности находятся части сегментальных органов двух соседних пар. Каждая трубочка извивается петлеобразно в своей средней части и состоит из трех отделов: открытой в полость тела мерцательной воронки, вытянутой на другом конце в мерцательный канал, следующего за последним железистого отдела и выводного канала, образующего пузыревидное расширение перед своим выводным отверстием. Если общая полость тела вытеснена паренхимой, воронки нефридиев могут замыкаться клетками, как в первом случае. Из нефридиев выводятся почки позвоночных, которые представляют собой группы нефридиев, впадающих с каждой стороны в особый выводной канал — мочеточник и связанных в одну компактную массу соединительной тканью. При этом воронки в огромном большинстве случаев зарастают, и жидкое выделение выводится не из общей полости тела, а из крови, приносимой в почки особыми кровеносными сосудами. У членистоногих почки представлены выше упомянутыми (ст. 255) Мальпигиевыми трубочками, которые в разном числе впадают, как выше упомянуто, в кишечный канал. И эти образования, выделяющие азотистые продукты в кристаллическом виде, могут быть выведены из сегментальных органов. По-видимому, сегментальные органы, находящиеся в передних сегментах тела, могут утрачивать свою связь со стенками тела, а через них и с наружной средой, и, не меняя своей железистой функции, становиться железами другой категории, которые являются придатками кишечного канала. В некоторых сегментах сегментальные органы совершенно недоразвиваются, в некоторых — упрощаются в строении и становятся просто выводными протоками половых органов.
7) Органы размножения, или половые органы, состоят из существенных и дополнительных частей. К первым относятся скопления эмбриональных клеток, известные под названием гонад, которые при раздельной полости образуют у самок яичники (женская гонада), у самцов — семенники (мужская гонада). В женской гонаде эмбриональные клетки развиваются в яйца, в мужской — в сперматозоиды (см. выше). Форма яичников, как и семенников, может быть весьма различна, и то это плотные органы, то трубчатые. Однако, и кажущиеся снаружи плотными гонады в действительности часто имеют внутреннее трубчатое строение. Дополнительные части половых органов являются их протоками и различными железистыми придатками. Протоки половых органов не всегда находятся в постоянной непрерывной связи с их существенными частями.
8) Нервная система является в ее простейшем виде сетью нервных клеток и их отростков, но уже у кишечнополостных во многих случаях позволяет видеть дифференцировку нервного кольца, которое в таком случае должно быть рассматриваемо в качестве центрального отдела нервной системы, тогда как остающиеся вне его, но связанные с ним нервные клетки и волокна образуют в таком случае периферический отдел нервной системы. Еще резче это деление на центральную и периферическую нервные системы выражено у иглокожих, где, как и у кишечнополостных, нервная система устроена по лучевому плану строения. Начиная с низших червей, центральная нервная система устроена по плану двубоковой симметрии и состоит либо из парного нервного узла в переднем конце тела либо из парного по внутреннему строению ствола, который лежит в срединной плоскости и произошел из двух парных сблизившихся до срастания стволов. Такой ствол образует в своем переднем конце сложное нервное кольцо, окружающее пищевод, а на дальнейшем протяжении несет посегментно расположенные нервные узлы. Путем сближения и слития этих узлов достигается укорачивание центральной нервной системы описанного строения. Этот тип нервной системы наблюдается у разных червей и членистоногих, причем у последних к центральной нервной системе с ее периферическими нервами присоединяется т. н. симпатическая нервная система, заведующая отправлениями органов пищеварения. У моллюсков устройство центральной нервной системы в основе сводится к трем парам нервных узлов, занимающих определенное положение в теле животного и соединенных друг с другом определенными комиссурами. Из слития этих узлов получается, в свою очередь, нервное кольцо вокруг пищевода, а из их расхождения, с развитием дополнительных нервных узлов, очень сложная нервная система, сложность которой увеличивается еще благодаря утрате моллюском двубоковой симметрии. У позвоночных центр, нервной системы состоит из головного и спинного мозга, периферическая — из отходящих от них нервов; симпатическая представлена парной цепью, лежащей по сторонам спинного мозга и связанной с нервными узлами, которые принадлежат основаниям спинномозговых нервов. Головной мозг в ряде позвоночных проходит постепенные осложнения, которые приводят его к чрезвычайной сложности, что выражается не столько во внешнем, сколько во внутреннем строении. Кроме того, у позвоночных вокруг головного и спинного мозга развивается определенная система нескольких оболочек, имеющих большое значение в деле охранения и питания центральной нервной системы. Периферические нервы делятся на чувствующие и двигательные, смотря по тому, соединяют ли они с центральной нервной системой органы чувств или идут от нее к мышцам. Многие нервы — смешанного характера, состоят из волокон двоякого рода, т. е. как чувствующих, так и двигательных.
9) Органы чувств представлены органами пяти специальных чувств — осязания, вкуса, обоняния, зрения и слуха — и т. н. боковой линией рыб и водных личинок амфибий, которая способна воспринимать механические и, вероятно, термические раздражения, будучи менее специализованным, более общим органом чувств. Иногда боковые линии называют органом шестого чувства, что неправильно, т. к. собственно 6-го чувства мы не знаем. Существенную часть всякого органа чувств составляют эпителиальные клетки, измененные особым образом.
Их поддерживают особые промежуточные клетки, а, кроме того, в связи с ними развиваются разные другие приспособления, иногда очень сложного строения. Но в простейшем виде чувствующие элементы связаны с общими покровами и являются чувствующими клетками, разбросанными среди обыкновенных эпителиальных клеток покровов. Из таких клеток берут начало органы осязания, вкуса и обоняния, путем локализации клеток чувств, эпителия на определенных местах, без ограничения свободной поверхностью тела; у позвоночных орган вкуса принадлежит ротовой полости, в органе обоняния клетки чувствующего эпителия также погружены на дно мешковидных впячиваний на переднем конце головы, которые у большинства заменяются носовыми ходами, вступая вторично в соединение с полостью глотки. У позвоночных осложнение органов обоняния заканчивается образованием т. н. носовых раковин с поддерживающим их скелетом. Органы зрения, или глаза, намечаются у реснитчатых червей пигментными скоплениями, стоящими в связи с нервным узлом. Затем, они осложняются путем присоединения преломляющего свет тела, образования чашковидного впячивания, которое выстлано зрительным эпителием, или сетчаткой, и либо остается открытым, либо закрыто т. н. линзой. Такого рода изменения глаз можно проследить во всех подробностях у разных червей и моллюсков, тогда как для членистоногих характерны т. н. сложные фасетированные глаза, состоящие из скопления большого числа бокалообразных простых глаз или глазков, каждый с своей собственной роговицей. Таким образом, роговица сложного глаза складывается из роговиц его составных частей, почему и получается фасетированная поверхность, в связи с чем изображение предмета, схватываемое таким глазом, является мозаичной картиной, т. е. каждая составная часть глаза видит только часть рассматриваемого предмета, а не весь предмет. Глаза позвоночных по внешнему виду представляют удивительное сходство с глазами головоногих моллюсков, но в действительности резко разнятся от них, и это сходство относится, таким образом, к явлениям конвергенции. Дело в том, что тогда как у моллюсков элементы сетчатки направлены в сторону, к источнику света, как у всех беспозвоночных, у позвоночных они обращены в противоположную сторону, и впечатления получаются не от видимого предмета непосредственно, а от изображения предмета, получающегося на экране позади сетчатки. Глаз позвоночных представляет собой глазное яблоко, помещающееся в полости глазницы и защищенное верхним и нижним, а у некоторых еще и третьим веком (мигательная перепонка). Глазное яблоко представляет собой полое образование с толстыми, спереди прозрачными (роговица) стенками, полость которого разделена линзой (хрусталик) и другими придаточными образованиями на передний, больший, и задний, меньший, отделы и выполнена в передней части т. н. водянистой влагой, в задней — стекловидным телом. Клетки чувствующего эпителия изменены в зрительные элементы сетчатки. К категории органов зрения относится также т. н. темянной глаз — глазоподобный непарный орган, соединенный с крышей 3-го желудочка головного мозга. По строению он сходен с глазами беспозвоночных и, по всей вероятности, у предков некоторых современных позвоночных функционировал в качестве глаза, хотя едва ли может считаться функционирующим органом у современных позвоночных даже там, где сравнительно мало изменен в смысле недоразвития. Наконец, орган слуха берет свое начало в т. н. слуховых пузырьках, которые существуют у медуз из кишечнополостных и у некоторых ракообразных, функционируя в качестве органов сохранения равновесия и ориентировки тела в пространстве. Несомненные органы слуха имеются у насекомых, но своего полного развития они достигают у позвоночных. Здесь они являются парным органом, срастающимся с черепом в его височной области, и состоят из существенных и дополнительных частей. Существенные части представлены перепончатым лабиринтом, который погружен в хрящ или кость височной области и состоит из полукружных каналов и улитки, из коих первые несут функцию органов для сохранения равновесия, тогда как собственно воспринимающий звуки аппарат лежит в улитке. Перепончатый лабиринт образует т. н. внутреннее ухо, и дальше этого развитие органа слуха у рыб не идет. Начиная с амфибий, к внутреннему уху присоединяется среднее, образующее барабанную полость, замкнутую снаружи т. н. барабанной перепонкой. Среднее ухо развивается из остатков верхней части жаберной щели, лежащей между челюстной и подъязычной дугой (брызгальце), и соединяется Евстахиевой трубой с глоткой. Наконец, у млекопитающих кнаружи от среднего уха лежит внешнее, состоящее из наружного слухового прохода, вход в который защищен ушной раковиной, с сложным хрящевым скелетом. Все органы чувств связаны с определенными нервами. Так, к органам обоняния идут нервы 1-ой пары головного мозга (обонятельные), к парным глазам — идут нервы 2-ой пары головного мозга (зрительные), к органам слуха — идут нервы 8-й пары головного мозга, и т. д.
Размножение и развитие сложных животных. Сложные животные размножаются бесполым и половым путем. Бесполое размножение представлено почкованием и делением и весьма распространено у кишечнополостных, у червей и оболочников. Однако, оно комбинируется здесь с половым размножением, т. е. из яйца происходит особь, которая уже затем размножается почкованием или делением, причем дает, таким образом, или отдельные особи или колонии из многих особей. Если размножение почкованием или делением сопровождается срастанием вновь получающихся особей, колония приобретает чрезвычайно запутанное строение, что, например, наблюдается у губок. У червей бесполое размножение приводит к образованию цепных форм. Половое размножение происходит из яйца, оплодотворенного (что должно считаться нормальным) или неоплодотворенного (что очень редко) сперматозоидом. Для огромного большинства животных нормой является разделение полов, т. е. существование самцов и самок, но в разных группах наблюдается правильно гермафродитизм (см.). Иногда самка откладывает яйца, и такие животные называются яйцеродными; иногда оплодотворенное яйцо дает начало зародышу еще внутри материнского тела, но зародыш не срастается с материнским телом и рождается в яйцевых оболочках, от которых освобождается при рождении, — такие животные называются яйцеживородными; наконец, у третьих зародыш, развивающийся внутри материнского тела, срастается с ним посредством особых придатков, питается на счет питательных соков матери и рождается свободным от оболочек, — такие животные называются живородными. Если размножение происходит без оплодотворения яиц, оно называется партеногенетическим. Некоторые животные размножаются в личиночном состоянии, и такое явление называется пэдогенезисом. У некоторых животных, например, сальп, половое и бесполое размножение с поразительной правильностью чередуются между собой, представляя т. н. перемежающееся размножение. При этом бесполое поколение представлено одиночными особями, которые размножаются почкованием и таким образом производят цепные формы; особи цепных форм являются половыми и размножаются яйцами. У разных кишечнополостных перемежающееся размножение очень распространено, и в некоторых случаях господствующим поколением является половое, в некоторых бесполое. Иногда чередуются два различных половых поколения, распознаваемые по внешним признакам.
У огромного большинства животных самцы и самки разнятся между собой по внешнему виду. У насекомых существует еще явление сезонного диморфизма, когда особи, выходящие из перезимовавших и летних куколок, более или менее сильно разнятся между собой. Но затем, существуют животные, именно общественные насекомые, у которых существуют не две, а три и даже четыре различные формы особей, т. е., кроме самцов и самок, существуют еще рабочие особи и воины. Но наибольшей сложности полиморфизм особей достигает у сифонофор, где, при чрезвычайной специализации функций, отдельные особи колонии становятся не отличимыми от органов.
Готовый к рождению зародыш иногда разнится от взрослого животных очень мало, иногда весьма существенно. В первом случае он доразвивается, главным образом, путем роста; во втором — он претерпевает ряд сложных и подчас резких изменений. Например, новорожденный кузнечик во всем существенном сходен со взрослым насекомым и становится таковым путем роста, после последовательного ряда линек. Такое развитие называется прямым, или развитием без превращения, без метаморфоза. С другой стороны, у бабочки из яйца выходит червеобразная личинка — гусеница, которая, выросши до известного предела, переходит в покоящуюся стадию — куколку. Под защитой оболочек куколки, одетой иногда т. н. коконом, формируется взрослое насекомое, и, спустя некоторое время — иногда несколько недель, иногда несколько месяцев — по разрыве покровов куколки, из последней выходит совершенно готовое насекомое. Такое развитие называется непрямым, или развитием с превращением, с метаморфозом. Примеры его имеются и у позвоночных, среди амфибий, где у лягв и жаб из яйца выходит хвостатая личинка — головастик, который, спустя то или другое время, быстро превращается в бесхвостую взрослую форму. Обыкновенно метаморфоз сопровождается частичным разрушением личиночных органов и построением из остатков их соответствующих органов взрослого животного. Это явление разрушения личиночных органов и тканей известно под названием гистолиза. С биологической точки зрения, животные могут быть разделены на свободно живущих, и паразитирующих. Должно считаться, безусловно доказанным, что паразиты развились из свободно живущих путем приспособления к своеобразным условиям питания, причем паразитизм обыкновенно сказывается в большем или меньшем недоразвитии нервной системы, органов чувств и органов движения и в соответствующем большем развитии органов размножения. Но, кроме того, обращает на себя внимание т. н. симбиоз, т. е. сожительство двух организмов, полезное для обоих. Степень близости при таком сожительстве зависит от очень различных обстоятельств, и проистекающая от него выгода чрезвычайно разнообразна, хотя, по-видимому, в большинстве случаев сводится к целям защиты, особенно у высших животных. Наконец, надо отметить, что симбиоз наблюдается не только между животных, но и у животных с растениями.
Классификация. Сумма сведений о животных выражается в их классификации, т. е. в группировке всех ныне известных животных, — как вымерших, так и современных, — по определенному принципу. Этим принципом является большая или меньшая степень сходства разных животных между собой, объясняемая, согласно эволюционному учению, единством происхождения всех животных и более или менее близким родством разных животных друг другу. Чтобы определить степень сходства или несходства разных животных, — во-1-х, сравнивают их строение во взрослом состоянии, во-2-х, изучают историю их развития. В некоторых случаях очень большую помощь при определении родственных отношений между животными оказывает палеонтология, с ее сведениями об ископаемых животных. При построении классификации исходят из изучения систематических единиц, называемых видами. Под видом разумеется собрание особей, похожих друг на друга настолько, насколько походят друг на друга дети одних и тех же родителей. Близкие виды группируют в роды, рассматриваемые в качестве членов различных семейств: семейства соединяются в отряды, отряды — в классы; наконец, классы соединяются в типы. Объем типов, как и других систематических групп, может быть чрезвычайно различен. Есть роды с одним только видом каждый; есть и семейства, содержащие только по одному роду. Даже некоторые типы чрезвычайно бедны группами, нижестоящими в системе, тогда как другие — чрезвычайно обширны. Чтобы судить об этом, достаточно сказать, что если принять число всех известных видов животных около 400 тысяч, одних насекомых известно около 250 тысяч, т. е. более половины всех животных, а насекомые составляют только один из классов типа членистоногих. Напротив, тип плеченогих представлен сравнительно немногими видами, вообще, и очень немногими в современной фауне. В настоящее время большинством зоологов принимаются следующие типы:
I. Простейшие животные, Protozoa, в огромном большинстве случаев — микроскопически малые организмы, являющиеся свободно живущими клетками. Одни из них одеты раковинкой; другие имеют скелет, представленный иглами; третьи совершенно лишены твердых образований в своем теле, которое всегда состоит из протоплазмы с ядром или ядрами. Несмотря на простоту строения простейших животных, жизненный цикл многих из них крайне сложен. Способы размножения простейших крайне разнообразны. Все остальные типы животного царства соединяются в большой отдел Metazoa, характеризуемый многоклеточностью относящихся сюда организмов и дифференцировкой у них тканей. Типы Metazoa следующие:
II. Губки, Porifera. Сидячие, водные Metazoa, за исключением одного семейства, живущие в море. Первичная форма — в виде бокала или цилиндра, стенки которого пронизаны многочисленными отверстиями, ведущими в большую полость внутри животных. Путем утолщения стенок, неполного деления и затем срастания многих особей, получаются сложные колониальные формы. Скелет представлен или волокнами рогоподобного вещества, или кремневыми, или известковыми иголочками. Размножение происходит как половым, так и бесполым путем.
III. Кишечнополостные, Coelenterata. Лучистые животные, с 2, 4 и 6 радиусами или кратным числом из 4 и 6. В теле находится только одна система полостей, которые все сообщаются друг с другом, заменяя собой пищеварительную полость, общую полость тела и сосудистую систему. Размножение как бесполое, так и половое, иногда с чередованием половых и бесполых поколений. Чрезвычайно распространена колониальность. Многие выделяют массивный скелет (кораллы).
IV. Плоские черви, Platyhelminthes. Двусторонне симметричные животные с мягким, обыкновенно сплющенным телом, без настоящей сегментации. Кишечный канал может быть или не быть, но если есть, то открывается наружу только одним отверстием и представляет собой, в сущности, единственную полость в теле этих животных Между общими покровами и кишечником нет такого промежутка, который можно было бы сравнить с общей полостью тела других животных Выделительная система представлена так называемой водно-сосудистой системой. Мужские и женские половые органы обыкновенно соединены в одной особи.
V. Круглые черви, Nemathelminthes. Тело удлиненное и цилиндрическое, слегка заостренное на обоих концах. Поверхность гладкая, блестящая, но без слизи. Кишечный канал есть. Полы, за немногими исключениями, раздельны. Развитие у большинства сопровождается превращением.
VI. Коловратки и близкие формы, Trochelminthes. Микроскопически малые животные, по большей части принадлежащие пресноводным бассейнам. Наиболее характерными особенностями их служат венчик ресничек на переднем конце тела и жевательный аппарат в переднем отделе кишечника. При внешнем сходстве некоторых коловраток с инфузориями, это — многоклеточные животные, с кишечным каналом, общей полостью тела, органами выделения, нервной системой, органами чувств и раздельнополостью.
VII. Моллюсковидные животные, Molluscoida. У большинства есть настоящая общая полость тела, выстланная эпителием, в которой кишечный канал подвешен посредством мезентерия или заменяющих мезентерий тяжей. На переднем конце тела имеется особый лопастной вырост со щупальцами, а над ртом, у большинства, — особый чувствующий вырост. Нервная система представлена одним или двумя узлами или нервным кольцом. Органы выделения сокращены.
VIII. Иглокожие, Echinodermata. Морские животные лучистого строения, с пятью основными лучами, но при всем том чрезвычайно разнообразного наружного вида. Стенки тела имеют наружный известковый скелет, иногда представленный прочным панцирем. Разные системы органов достигают сравнительно высокой сложности. Так называемая водно-сосудистая система играет роль органов передвижения. Развитие — с превращением.
IX. Кольчатые, Annelida. Тело животных, соединяемых в этом типе, состоит из ряда следующих друг за другом сегментов, или метамеров, т. е. частей, похожих друг на друга. Поэтому таких животных называют сегментированными, а свойственное им деление тела на сегменты — сегментацией. Обыкновенно сегменты до крайности похожи друг на друга, но некоторые отличаются между собой, вследствие вторичных изменений. Обыкновенно у кольчатых есть большая общая полость тела, и у большинства система кровеносных сосудов. Нервная система состоит из головного узла, окологлоточных комиссур и парной брюшной цепи, сегментированной на ряд нервных узлов. Органы выделения представлены метамерно расположенными парами трубочек, нефридиями или сегментальными органами, которые ведут из общей полости тела наружу. Нервная цепь лежит на брюшной стороне; главный кровеносный сосуд — на спинной.
Х. Членистоногие, Arthropoda. Сходны с кольчатыми по метамерной сегментации своего тела, при его более или менее полной двусторонней симметрии, при положении рта и заднепроходного отверстия на противоположных концах удлиненного тела и при нервной системе из отдельно лежащего надглоточного узла (головной мозг) и брюшной, по положению, парной цепи ганглиев. Отличие, главным образом, заключается в том, что каждый неизмененный сегмент несет пару расчлененных придатков. Общая полость тела заменена кровеносными синусами, свободно сообщающимися с кровеносной системой. Отдельные сегменты обнаруживают наклонность сливаться по группам. Сердце лежит на спинной стороне.
XI. Слизняки, или мягкотелые, Mollusca. Мягкотелые получили свое название за то, что внутри их тела у них совсем нет таких твердых образований, какие есть у позвоночных. Хотя эта мягкость внутренних органов выражена, например, у членистоногих, но последние имеют твердые наружные покровы, определяющие собой очертания их тела, или т. н. наружный скелет. У слизняков также часто бывает наружный скелет, образованный преимущественно известковым веществом, но он прикрепляется только к части поверхности тела животных, обыкновенно очень толст и легко отрывается, почему его часто описывают самостоятельно, в качестве раковины. Эта раковина выделяется частью кожи, которая обыкновенно свешивается в виде лопасти, называемой мантией, и прикрывает в большей или меньшей степени тело животного, оставляя между, ним и собой промежуток, называемый полостью мантии. В этой полости находятся органы дыхания (жабры); сюда открывается почка или почки, заднепроходное отверстие и часто также — половые протоки. Будучи в типе двустороннесимметричными животными, слизняки во взрослом состоянии часто утрачивают это. Нервная система в простейшем виде представлена тремя парами нервных узлов, из которых окологлоточная соединена как с кишечной парой, так и с ножной, но последние первоначально не соединяются между собой. Органом передвижения является особый придаток тела — т. н. нога.
XII. Хордальные, Chordata. Тип хордальных животных обнимает собой весьма разнообразных по наружному виду и величине животных, связываемых следующими признаками: у всех у них или в течение всей жизни или на известных стадиях развития существует особый эластический тяж, т. н. спинная струна, или хорда, совпадающая по положению с продольной осью тела животного. Кроме того, на известной стадии развития у них у всех на стенках передней части кишечного канала есть щели, первоначально служащие для проведения воды в дыхательных целях и называемые жаберными щелями. Наконец, нервная система закладывается в виде полосы эктодермы, т. н. медуллярной полоски, которая, углубляясь, превращается последовательно в борозду, канал и замыкается в трубку. Этот тип разделяется на четыре подтипа.
Подтип 1. Hemichordata. Червеобразные, морские животные, с короткой зачаточной хордой. Известны под сборным именем баланоглосса.
Подтип 2. Cephalochordata. Маленькие рыбоподобные морские животные, известные под сборным именем ланцетника (Amphioxus). Спинная струна идет от одного конца тела до другого, над кишечником. Сердца нет. В центральной нервной системе едва дифференцирован зачаток головного мозга. Органы выделения представлены многочисленными нефридиями в жаберной области. Из оплодотворенного яйца развивается мерцательная личинка с продолжительным нарушением двусторонней симметрии во время ее развития.
Подтип 3. Tunicata, или Urochordata. Эта группа называется оболочниками — за своеобразную прочную, иногда толстую и периодически сбрасываемую, оболочку (туника). Во взрослом состоянии хорды нет, но в личиночном состоянии хвост, являющийся характерным органом передвижения личинки, поддерживается хорошо развитой хордой. Развитие происходит с превращением. Во взрослом состоянии, вследствие процессов регресса, организация чрезвычайно упрощается. Сюда относятся как простые, так и колониальные животные, — иногда с перемежающимся размножением, то сидящие, то плавающие, известные под названием асцидий, сальп и пр.
Подтип 4. Vertebrata. Последний подтип составляют позвоночные, названные так за то, что спинная струна у них отчасти или вполне заменяется сериально расположенными позвонками, которые, вместе взятые, образуют осевой скелет, называемый позвоночным столбом. Скелет головы образует череп, в котором заключен головной мозг. Конечностей никогда не бывает более двух пар. Челюстной аппарат образуется из измененной передней жаберной дужки. Всегда есть сердце того или другого строения. Органы выделения представлены почками, которые развиваются из нефридиев. Сюда относятся все рыбы, земноводные, или амфибии, пресмыкающиеся, или чешуйчатые гады, птицы и млекопитающие, — вместе с человеком.
М. Мензбир.
Объяснения к таблицам
Животные.
Таблица 1.
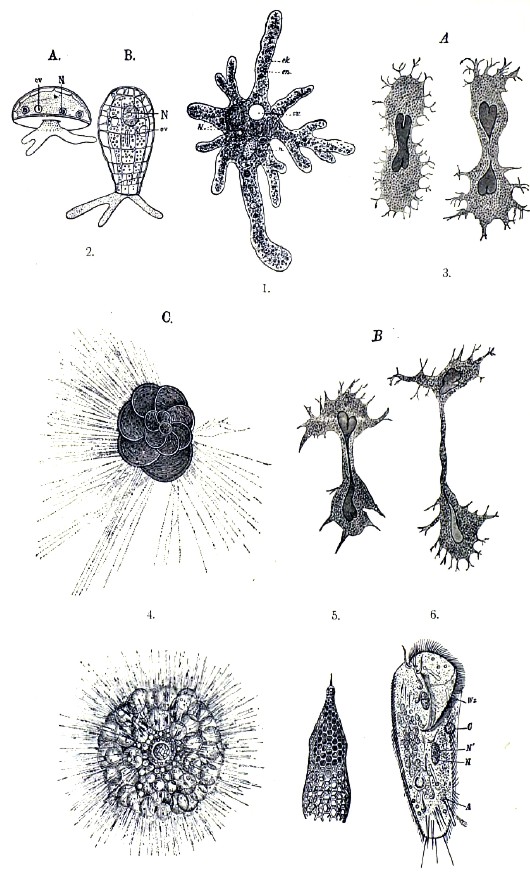
Рис. 1. Amoeba proteus. n — ядро; сv — сократимая вакуоль; N — пищевые скопления; еn — внутренняя зернистая плазма; ek -наружная однородная плазма.
Рис 2. Корненожки. А — Arcella vulgaris; В — Quadrula symmetrica; С—Rotalia venata. А и В с хитиновой раковиной и С — с известковой многокамерной.
Рис. 3. А и В — амебообразные белые кровяные тельца, на разных стадиях деления.
Рис. 4. Thalassicolla pelagica — радиолярия без раковины.
Рис. 5. Eucyrtidium lagena — радиолярия с колоколовидной раковиной.
Рис. 6. Stylonychia mytilus — инфузория. Ws — приротовой венчик ресничек; C — сократимая вакуоль; N — большое ядро; N’ — малое ядро; А — порошица.
Таблица 2.
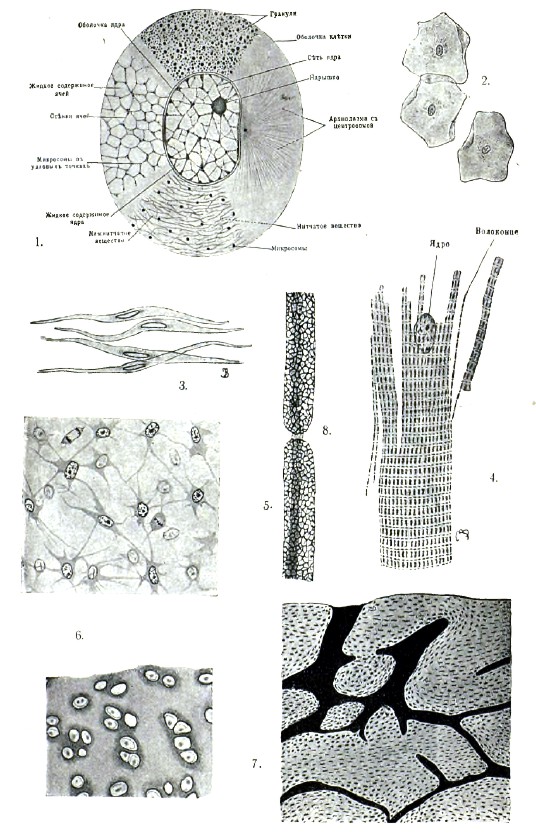
Рис. 1. Схематическое изображение клетки, на котором, в разных отрезках клетки, показана ее структура по Альтману (верхний отрезок — грапули), Бючли (левый — пенистое строение) и Флеммингу (нижний — нитчатое строение).
Рис. 2. Эпителиальные клетки.
Рис. 3. Гладкие мышечные волокна (мышечные клетки).
Рис. 4. Поперечнополосатое мышечное волокно.
Рис. 5. Эмбриональная соединительная ткань.
Рис. 6. Гиалиновый хрящ.
Рис. 7. Продольный шлиф трубчатой кости. Видны Гаверсовы каналы в расположение костных телец по отношению к ним и к поверхности кости.
Рис. 8. Сеть нейрокератина вокруг осевого цилиндра.
Таблица 3.
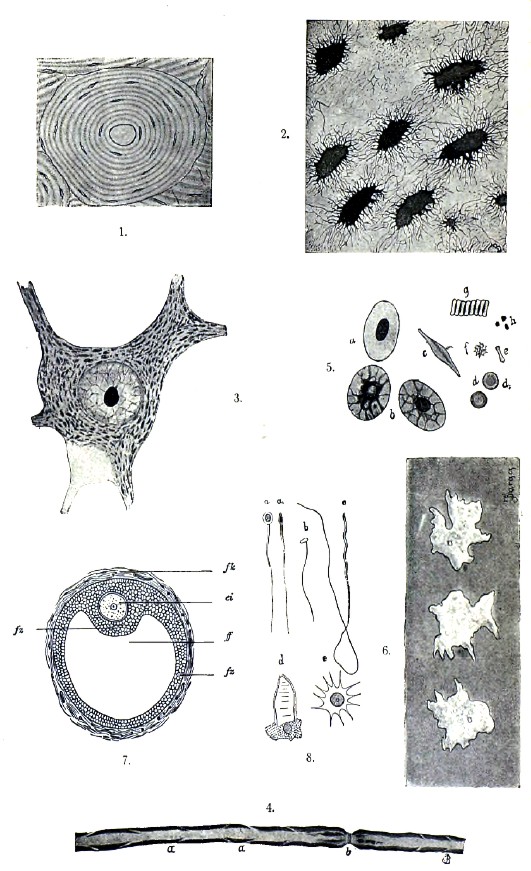
Рис. 1. Поперечный шлиф трубчатой кости.
Рис. 2. Костные тельца (полости), инъецированные краской, чтобы видеть форму костных клеток, их ветвление и способ соединения.
Рис. 3. Нервная клетка с ядром и ядрышком, увеличена около 950 раз.
Рис. 4. Нервное волокно с мякотным влагалищем. а — шмидт-лантермаповские пасечки, b — перетяжка Ранвье.
Рис. 5. а—с — красные кровяные тельца (эритроциты) лягушки; d—g — то же человека; h — кровяные пластинки.
Рис. 6. Белые кровяные тельца (лейкоциты) лягушки.
Рис. 7. Граафов пузырек из яичка млекопитающего с большим скоплением жидкости внутри. вi — яйцо: fs — окружающие яйцо и выстилающие стенки пузырька клетки; ff — жидкость в полости пузырька; fk — оболочка пузырька.
Рис. 8. Сперматозоиды. а, а1, — человека (прямо и в профиль); b — губки; с — рыбы (ската); d — круглого червя; в — краба.
Таблица 4.
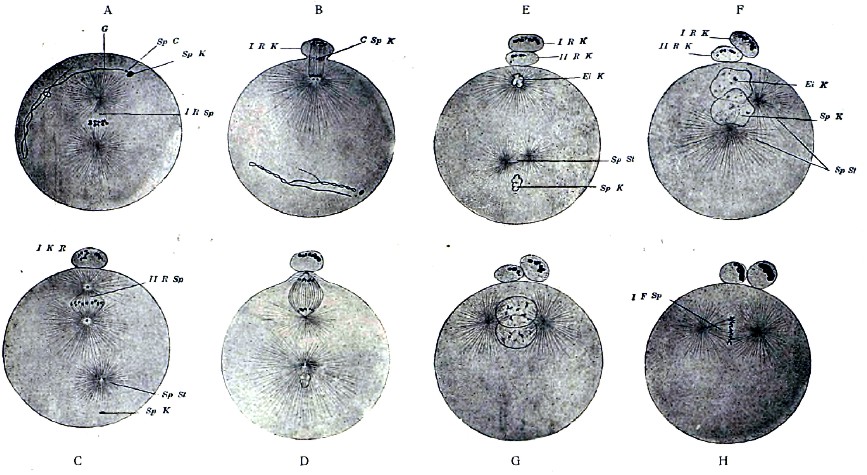
А—Н. Последовательные стадии оплодотворения яйца Physa fontinalis, по Костанецкому и Виржейскому.
А. Начало образования 1-го направляющего пузырька; сперматозоид весь виден в яйце.
В. Отделение первого направляющего пузырька.
С. Начало образования 2-го направляющего пузырька; деление центросомы сперматозоида с развитием вокруг нее лучистости.
D. Продолжение процесса; ядро сперматозоида становится пузыревидным.
Е. Отделение направляющих пузырьков закончилось; вокруг яйцевого ядра остатки лучистости.
F. Сближение ядра яйца и ядра сперматозоида.
G. Ядро яйца сходится с ядром сперматозоида; центросомы (сперматозоида) лежат по бокам их.
Н. Хромосомы двух ядер образуют звезду. С Sp К — центральное веретено; Еi К — ядро яйца; I F Sp — веретено 1-го деления яйца; G—хвостик сперматозоида; I R К — 1-й направляющий пузырек; IІ R К — 2-й направляющий пузырек; I R Sp — веретено 1-го направляющего пузырька; II R Sp — веретено 2-го; Sp С — центросома сперматозоида; Sp К — ядро сперматозоида; Sp St – лучистость сперматозоида.
Таблица 5.
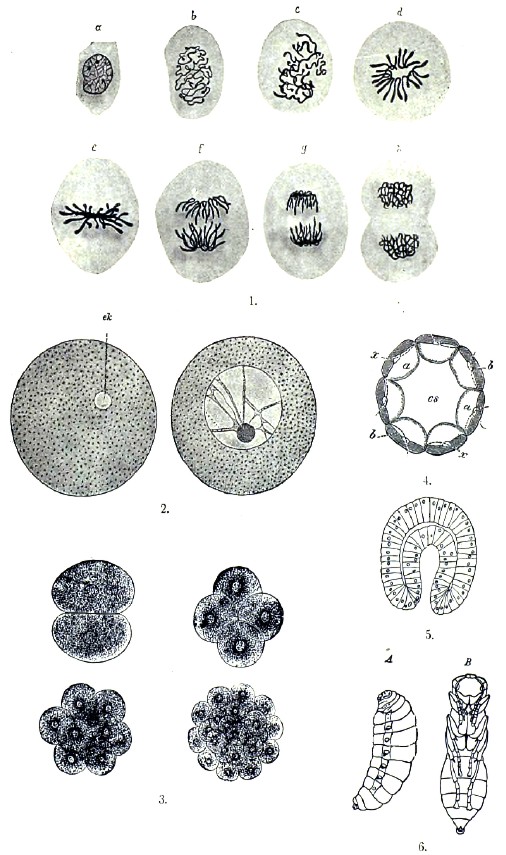
Рис. 1. Деление ядра в эпителиальных клетках роговицы головастика, а—покоящееся ядро, b, с—клубкообразная стадия, d - монастер, е — то же сбоку, f — диастер, g — ядра анафазы, h - восстанавливающиеся ядра дочерних клеток.
Рис. 2. Слева зрелое, справа незрелое яйца иглокожего.
Рис. 3. Последовательные стадии дробления лица.
Рис. 4. Бластула Gеryonia. С1 - бластоцель, а — внутренняя прозрачная плазма, b - наружная темная плазма; пунктиром намечены границы ближайших делений.
Рис. 5. Гаструла голотурии.
Рис. 6. Личинка (А) и куколка (В) осы.
Таблица 6.
Рис. 1. Гидра, два экземпляра, один с расправленными, другой с вытянутыми щупальцами; первый размножается почками. bd1, bd2, bd3 — почки на разных стадиях развития; t — щупальца; hyp — рот, а — постороннее тело.
Рис. 2. Гидроидный полип Bougainvillea ramcsa. А — целая древовидная колония (ест. вел.); В — веточка колонии, увеличена; С — медуза, hyd — гидранты, med — развивающиеся медузы.
Рис. 3. Губка Sycon gelatinosum, снаружи и в вертикальном разрезе, iр — вводящие поры; о - выводные отверстия (oscula).
Рис. 4. Трубчатый кольчатый червь Serpula.
Рис. 5. Брахиопода Magellania flavescens. А — целая раковина со спинной стороны, В — то же сбоку, d.v. — свинная, v. v. — брюшная раковина; b — вырост последней uf — его отверстие.
Рис. 6. Термиты, а — рабочий, b —воин, с — бескрылая самка, d — самка, готовая к размножению, в —крылатая форма самки или самца.
Таблица 7.
Рис. 1. Схематическое изображение внутренних органов скорпиона сбоку, аn. — заднепроходное отв.; ant. art. передняя аорта; brn. головной мозг; chel. chelicerae; hep. du. печеночный проток; hvt. сердце; lat. eye. боковые глаза; leg. 1—4, ножки; mal — Мальпигиевы трубочки; med. eye. средний глаз; mesent. — mesenteron; nе. со-нервная цепь; pect. гребневидные придатки; pedi.— pedipalpus; pois. gld. ядовитая железа; post. art. задняя аорта; proct. — proctodaeum; pul.1—pul.4 легочные мешки; sub. gang, подглоточный ганглий.
Рис. 2. Рак, Astacus fluviatilis, вскрытый с правой стороны, аа. артерия усиков; ab. брюшко; аn. заднепроходное отверстие; bd. желчный проток; bf. 4 клешня; bm. брюшная нервная цепь; cs. кардиальный отдел желудка; cth. головогрудь; cm. спинные мышцы; fm. брюшные мышцы; g. головной мозг; h. сердце; hd. толстая кишка; lr. печень; md. тонкая кишка; о. — ostium; оа. глазная артерия; oаа. верхняя брюшная артерия; ое. пищевод; рl. 1—5 плеоподы; рl. 6 уропод; ps. пилорический отдел желудка; sa. стернальная артерия; t. семенник и telson; uаа. нижняя брюшная артерия; vd. — vas deferens; vdo. мужское половое отверстие.
Рис. 3. Рак, Astacus fluviatilis, самец, сбоку, а1, anteunula; а2. antenna; ab. брюшко (шейка); cth. головогрудь; kd. жаберная крышка; r. — rostrum; 8. ногочелюсть 3-ей пары; 9. ножка 1-й пары; 10—13. остальные ножки; 19. uropod’ъ; XIV. первый сегмент брюшка; XIX. шестой сегмент брюшка.
Рис. 4. Органы кровообращения рака, схематично. Сосуды с артериальной кровью светлые, с венозной темные; стрелки указывают направление потока крови. g.жабры, h.сердце, р.околосердечная сумка (перикардий), v. венозный синус.
Рис. 5. Расположение главных стволов трахейной системы насекомого.
Таблица 8.
Рис. 1. Морская звезда с брюшной стороны, с вытянутыми амбулякральными ножками.
Рис. 2. Нервная система насекомых. А — муравья, В — майского жука, С — мухи, h — надглоточный, u — подглоточный нервный узел; sр — место прохождения пищевода. 1—3 — торакальные нервные узлы; а1—а3 — брюшные нервные узлы.
Рис. 3. Самец берегового слизня (lltorina), рис. 4 — самка его же. Животное нарисовано сверху, по извлечении из раковины, с отрезанной и отвернутой влево мантией, а — заднепроходное отверстие; d — слизистая желудка; f — нога; g — орган обоняния; gе — половое отверстие; h — сердце; k — жабра; l — печень; m — край мантии: mа — желудок; n — почка; n1 — вывод отв. почки; od — яйцевод: оv – яичник — копулят. орг.; r — семенная бороздка; s — семепровод; sd —слюнные железы; sp — пищевод; t — семенник; v — предсердие.
Рис. 5. Пластинчатожаберный моллюск, Anodonta cygnea, в разрезе, с левой стороны, а. заднепроходное отверстие; а ad. передний adductor; а. аv. передняя аорта; а. v. ар. отверстие auriculoventricul’apнoe; bl. мочевой пузырь; с. pl. gn. — cerebro-рleural’ный нервный узел; d. d. проток пищеварительной железы; d. gl. пищеварительная железа; d. р. а. спинное отверстие мантии; ex. sph. выводящий сифон; ft. нога; g. ар. половое отверстие; gon. гонада; gul. пищевод; i. l. j. мостики между листками жабр; in. sph. вводящий сифон; int. кишка; kd. почка; m. мантия; mth. рот; р. ао. задняя аорта; р. ad. задний adductor; рс. перикардий; pd. gn. peda’ный узел; r. ар. отв. почки; r. an. правое предсердие; rct. — rectum; r. р. а. отв. почки в перикардии; st. желудок; ty. - typhlosolis; v. желудочек сердца; v. gn. visceral'ный узел; w. t. водяные трубочки.
Таблица 9.
Рис. 1. Скелет форели.
Рис. 2. Внутренности лягушки в продольном разрезе, схематично; а — желудок; v — мочевой пузырь; с — тонкая кишка; сl — отв. клоаки; d — толстая кишка; е — печень; f — желчный проток; g — желчный пузырь; h — селезенка; t — легкое; k — laryux; l — жировое тело; m — семенник; n — мочеточник; о — почка; р — поджелудочная железа; s — главный мозг (полушария); sp — спинной мозг; t — язык; u — предсердие; ur — уростиль; v — желудочек сердца; vs — семенной пузырь; w — зрительная доля (головного мозга); х — мозжечок; у — отв. Евстахиевой трубы; s — носовой ход.
Рис. 3. Асцидия, схема продольного разреза, с левой стороны, после удаления оболочки и мантии. atr саv. атриальная или клоачная полость; atr. siph. атриальный сифон; br. car. — arteria branchlo-cardiaca; card. visс. — art. cardio-visceralis; gonod. половой проток; ht. сердце; hyp. гипофиза; mant. мантия; n. gn. нервный узел; oes. пищевод; оv. яичник; rect.—rectum; stig. stigmata; stom. желудок; tent. щупальца; test. оболочка; tr. v. поперечный сосуд; vent. v. брюшной сосуд; visc. br.—art. viscero-branchialis.
Рис. 4. Послойно вскрытая асцидия, с правой стороны. Большая часть оболочки и мантии с правой стороны снята, чтобы показать отношение этих образований к внутренним полостям, кишечному каналу и пр. an. заднепроходное отверстие; atr. ар. отверстие клоаки; end. эндостиль; gon. половые органы (гонады): gonod. их выводной проток; hyp. гипофиза; hyp. d. канал гипофизы; mant. мантия; nе. gn. первый узел; oes. ар. вход в пищевод; or. ар. ротовое отверстие; ph. глотка; stom. желудок; tent. щупальца; test. оболочка.
Рис. 5. Схема превращения хвостатой личинки в сидячую асцидию. А, стадия свободноплавающей личинки; В, только что прикрепившаяся личинка; С, более поздняя стадия; adh. сосочки прикрепления, atr. клоачная полость; cil. gr. мерцательная ямка; end. эндостиль; ht. сердце; med. туловищный нервный узел n. gn. нервный узел; noto. хорда; or. ротовое отверстие; red. — rectum; sens. ves. чувствующий пузырек; stig. — stigmata; stol. столон; t. хвост.
| Номер тома | 20 |
| Номер (-а) страницы | 210 |